Геномузыка на тринадцатой хромосоме
Для биоматематиков
 Проведя исследование заявления инопланетян, опубликованного в газете “Совершенно секретно“ (№№54, 55, 1999 и др.), и обобщая результат, примерно в том виде, как это сделано на сайте https://www.pravda.ru/science/ , автор высказал соображения относительно его фабрикации как фальшивки. 25.6.01 он направил соответствующий запрос в редакцию этой газеты. Прошло три года, редакция отмолчалась.
Проведя исследование заявления инопланетян, опубликованного в газете “Совершенно секретно“ (№№54, 55, 1999 и др.), и обобщая результат, примерно в том виде, как это сделано на сайте https://www.pravda.ru/science/ , автор высказал соображения относительно его фабрикации как фальшивки. 25.6.01 он направил соответствующий запрос в редакцию этой газеты. Прошло три года, редакция отмолчалась.
Можно делать множество предположений о причине молчания уважаемой газеты, но одна из них (как наиболее простая) могла крыться и в том, что редакция имеет для этого веские причины. Это побудило автора продолжить работу в поисках ответа на вопрос о земных технологиях инопланетян, если их посещения были реальны. Некоторые предположения о них, он доводит до читателя.
ЗЕМНЫЕ ТЕХНОЛОГИИ ИНОПЛАНЕТНЫХ ВИЗИТЕРОВ
Профессор музыки Дэвид Хьюрон из университета Огайо, предполагает, что музыка может быть эволюционным приспособлением, подобным прямохождению и речи, и позволившим человеку пройти весь путь эволюции (в смысле Дарвина). Он и его сторонники отстаивают точку зрения, указывающую на ее биологические корни, утверждая, что музыка возникла вместе с видом homo sapiens задолго до того, как особи этого вида расселились по всей земле, и было это примерно 40 тысяч лет тому назад. Об этом свидетельствуют, как говорит профессор, и археологические находки.
На недавнем конгрессе Нью-йоркской академии наук сотрудница Монреальского университета Изабель Перетц рассказала об исследованиях, подтвердивших наличие в мозге человека области, “ответственной” за музыку. Это, по ее мнению, доказывает, что “музыкальный дар” человека развивался в ходе эволюции, из чего следует, что в геноме человека должен быть “специальный ген музыки”. Сторонники такой “эволюционной теории” музыки считают, что музыкальный ген появился у человека десятки, а то и сотни тысяч лет назад, и он “в ходе естественного отбора должен был дать его обладателю большие возможности для размножения и распространения”. Более того, Дэвид Хьюрон полагает, что существует не один, а много генов, определяющих музыкальные способности человека. Сандра Трегуб из университета Торонто, также утверждает, что музыка это не изобретение человека, и что она возникла как инстинктивная форма отношений “мать-детеныш” и имеет приспособительное значение в коммуникационном обмене матери и ребенка.
Она обосновывает предположение, что генетическая предрасположенность к музыке возникла как инструмент воздействия на детеныша, способствующий сохранению вида, ибо мелодия помогала успокоить его и тем самым избежать опасности от издаваемых им криков и плача, которые могли привлекать блуждающих в поисках пищи хищников. И еще один довод ее сторонников: музыку используют птицы и животные не только для успокоения птенцов и детенышей, но и для привлечения партнера. Но животные, не обладая творческими возможностями, не могут “придумать” музыку, создать мелодию. Значит, она дана им природой, и это подтверждает ее генное происхождение. Добавим: это подтверждается и тем, что животные и птицы воспринимают музыку как определенный музыкальный строй и адекватно реагируют на него.
В июне 2001 года средства массовой информации сообщили об открытии японских ученых-генетиков, утверждающих, что код ДНК человека представим в виде мелодии, а гены, из которых он состоит, в виде нот. Сообщение иллюстрировалось показом графического отображения кодов генной мелодии, и сопровождалось ее озвучиванием. Свое заявление они подтвердили демонстрацией записи такой мелодии (мы ее будем называть “геномузыкой”), в виде потока цифровых кодов, которые были переложены на ноты, а затем и озвучены. Геномузыка, говорят японские ученые, отражает структуру и состав ДНК человека.
Концепция генного происхождения упорядоченного потока звуков, не обязательно в музыкальном строе, но несущих генную информацию, как это ни фантастично звучит, тесно связана с интригующими загадками посещения нашей планеты визитерами из других миров. Стараясь преодолеть косные стереотипы собственного мышления, рассмотрим эту загадочную проблему.
В исследованиях, направленных на изучение механизмов вероятной коррекции процессов естественной эволюции внеземными пришельцами (ВВ), мы пришли к результатам, указывающим на возможность внесения в структуры управляющих систем организма, новых управляющих механизмов. В частности, это механизм сохранения, как составная часть некоего биологического гомеостата – инструмента, обеспечивающего мультиустойчивость биологических процессов к внешним воздействиям. Такой инструмент призван играть важную роль собственного средства внутривидового отбора. Появились обоснованные аргументы считать, что это средство могло быть встроенным в распределенную биологическую компьютерную систему организма, в виде целевой программы на генном уровне, направленной на замену функции сохранения жизнедеятельности – гомеостазиса, на антагонистическую ей, функцию сохранения разрушения - антигомеостазиса.
Однако результаты исследования, подтвердив такую гипотетическую возможность, не позволили высказать какие либо соображения об инструменте ее реализации - механизме доступа на генный уровень. Это вносило серьезную неопределенность. В самом деле, сомневались мы, каким образом “технически” можно вносить воздействиях на генном уровне на малочисленных популяциях прародителя человека, разбросанных на громадном пространстве планеты? Просто утверждение, что визитеры знали, “как это делать”, порождая неуверенность, ничего не доказывает. Открытие геномузыки позволяет высказать рабочую гипотезу, частично снижающую такую неопределенность.
Гипотетическое воздействие коррекции, должно задавать целевую функцию эволюции носителю генетической информации организма - ДНК, высокополимерному соединению, содержащемуся в ядрах клеток и образующему, вместе с белками гистонами, вещество хромосом. В свете наших исследований, представимость кода ДНК цифровыми последовательностями акустических сигналов, дает основание предположить существование средств непосредственного доступа на уровень генома, а также механизма внесения, средствами этого доступа воздействий, корректирующих динамику эволюционного процесса.
Мы предположили, что структура управляющей цифровой последовательности, имея характер потока посылок – пачек, несущих закодированную в кодах генома информацию о стратегии корректирующего воздействия, имеет выделенную адресную часть – первую посылку, образующую передний фронт пачки. Изучая характер вносимых информационных потоков, не имея возможностей (и не ощущая необходимости) представления в явном виде стратегии управлений, транслируемых пачками импульсов, мы предполагали, что они коммутируют с естественными операторами управления эволюционным процессом. При этом, операторы управления отображались уравнениями в абстрактном виде, отношения взаимодействий переходных процессов на популяциях, графами - в лексической форме.
Если код ДНК человека, как носителя генетической информации, представим в виде мелодии, как это утверждают ученые-генетики, а определенные гены, соответствующие ее участкам, отображаются в нотном коде, то это может означать, что акустические колебания, полученные в результате демодуляции цифровой последовательности генетической информации, являются ее физическим носителем. Если это так, то такие колебания реализуют носитель не просто информации, но информации непосредственного доступа в системе информационного обмена “ДНК-объект внешней среды”. Под объектом внешней среды (ОВС), во избежание коллизий, необходимо понимать любой объект, внешний относительно молекулы ДНК, способный воспринимать генную информацию, независимо от собственного генетического базиса, ибо мы не можем делать не только предположения о характере генной системы визитеров, но даже и о самом ее существовании. Если верить исследованиям японских ученых, человек в статусе ОВС в состоянии распознать генную информацию собственного биологического вида, используя технические средства. Мы, в рамках гиотезы, можем полагать, что внеземные визитеры средствами распознавания генной информации ДНК прародителя человеческого рода, а также и клонируемого ими homo sapiens – органными или техническими, владели.
К сожалению, моделирование не позволило получить полного разрешения еще одного интересного вопроса – существовала ли естественная система доступа к генной информации клонируемого прародителя до первого вмешательства ВВ в процесс эволюции или она появилась как один из результатов их творческих усилий. Тем не менее, некоторые логические предпосылки, порожденные свойствами дискрет потока вносимых ВВ управлений и отраженные некоторыми особенностями моделей, допускающие такие предположения, существуют. По крайней мере - для первичного формирования множества адресов доступа.
Говоря о процедуре реализации прямого доступа в системе коммуникационного взаимодействия с ДНК, мы имеем в виду партнера, способного не только воспринимать генетическую информацию ДНК в виде “геномузыки”, так как это продемонстрировали японские ученые, но и реализовать обратный обмен так, возможно , как это могли делать внеземные визитеры. Это означает, что мы приписываем такому партнеру способность вносить генетическую информацию в геном человека извне, в виде потока должным образом модулированных цифровых кодов - той же геномузыки (но музыкального строя, соответствующего вносимой информации), воспринимаемой адресатом – геномом ДНК, ответственным за ее исполнение - рецепцию.
В исследованиях реальности сообщения ВВ, мы полагали, опираясь на сообщенные ими сведения о количестве посещений нашей планеты, что расписание, как временнáя стратегия внесения управляющей информации на интервалах времени посещения, имеет непрерывно-дискретный и регулярный характер. При этом, календарно-временные параметры вмешательства в ньютоновской системе отсчета времени, определяются на интервалах дискретности между пачками дискрет управления и длительностью во времени этих пачек.
Принималось, что количество таких пачек сигналов соответствует числу посещений (оно подлежало определению), некоторое число пачек образует цикл, и что собственно управляющая информация модулировалась генными кодами внутри каждой пачки. Считалось также, что непроизводительных посещений не было, а задачи каждого из них были выполнены. Интервалы дискретности между циклами рассматривались как интервалы переходных процессов рецепции.
Такое предположение (одна пачка сигналов коррекции на посещение и одна дискрета на цикл процессов рецепции), может быть обосновано тем, что расписание управлений могло быть заложено непосредственно в оператор (стратегию) управления. При этом отразилась возможность, для визитеров, с одной стороны, задавать время, потребное для реализации инерционных переходных процессов рецепции на популяциях вида, с другой – обеспечить расписание контрольно-корректирующих посещений планеты Земля, которые, надо полагать, также требовали от ретикулян определенных усилий.
Уточним, что под переходным процессом мы понимали ситуации, возникающие в “текущем” времени, как последствия рецепции управления на популяциях, отображающие совокупные изменения собственно воспринимающей системы - генома, и, как следствие, множества динамических отношений, образующих внутривидовые перетоки множеств (из предыдущего подвида в следующий) на популяциях. При этом мы полагали, что изменения генома, как воспринимающей системы, реализуются быстротечной (по отношению к инерционным физиологическим изменениям на популяциях) перекодировкой собственной системы кодирования и потребное на это время включалось в общее время рецепции.
Это означает, что моделирование отображало как изменение отношений на генном уровне управления, так и рецепцию воздействия на уровне множеств элементов популяции, подвергавшихся воздействию.
Интересно отметить, что основным механизмом рецепции на множествах популяций вида, как показал численный анализ эволюционных изменений геометрии черепа вида homo sapiens (примерно за 14 млн. лет), могло быть сохранение многообразий: корректируемая эволюция реализовалась сохранением функций. С точки зрения Дарвина - парадокс: эволюция через сохранение. Но это – самостоятельный элемент исследования, во многом подтверждающий правоту утверждений Ламарка. Ретикуляне знали, что делали!
Не имея каких либо оснований для предположений о характере и уровнях системы контроля рецепции, используя свободу логического выбора, мы полагали, что визитеры при своих посещениях, осуществляли прямой доступ к генной информации и могли производить косвенную оценку текущего состояния популяции по изменениям состава и структуры ДНК, относительно заданных ими на предыдущих циклах вмешательства. Считалось, что эти оценки они использовали как показатели качества рецепции для выработки последующих управлений по принципу “внешней” обратной связи. Мы моделировали отношения такой связи и объекта управления, коммутируя блоки коррекции, вводимой ВВ, с ключевыми блоками модели естественного эволюционного процесса, что превращало ее в модель процесса, управляемого извне. Такая блок-схема, настроенная на вид homo sapiens, была заложена в граф-октаву. как в модель третьего уровня моделирования. На этой модели, в терминах лексики, отображалась гипотетическая система отношений рецепции на популяциях, определенная блок-схемой управления: наше исследование осуществлялось как мультимодельное.
Мы используем, в этой статье, прошедшее время. Однако анализ показывает, что для завершения переходных процессов полной спирали последнего этапа коррекции эволюционного процесса, потребуется еще один ее цикл, состоящий, как и предыдущие, из 5 витков, что потребует от визитеров еще 5 посещений нашей планеты. Не для контроля ли достигнутого качества рецепции в 13-м цикле коррекции и закрепления ее результата в заключительном, 14-м цикле, как фиксации начальных условий последующей спирали эволюции, они совершили (если совершили) свое 65-е посещение?
Представление эволюционного процесса с обратными связями в цепи управления, при попытке оценки календарных сведений о посещениях, отобразило существование отношения масштабности между “реальным” временем ньютоновской модели, в котором отображаются витки эволюции, и биологическим временем, в котором отображаются витки ее коррекции, как результата рецепции. Оказалось, что витки коррекции отображаются в трехмерном ньютоновом временном пространстве эволюции спиралью, внешней относительно ее витков, как бы “навитой” на нее.
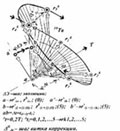
Эта спираль (показана на рисунке), характеризуется неравномерным и случайным во времени рецепции шагом ее закрутки, плоскость которой непрерывно изменяет свое угловое положение в пространстве, одновременно перемещаясь вдоль оси r витка эволюции со случайной скоростью шага закрутки. Радиус-вектор спирали, также случайной величины, вращается в переносном движении плоскости закрутки в направлении развития эволюционного процесса. Навитая спираль коррекции в явном виде отражает свое “внешнее“ относительно спирали эволюции происхождение, и такой же характер “производственного вмешательства” в ее естественный процесс.
Логично предположить, что случайность значений множества элементов атомарных “образующих” геометрии спирали коррекции, отражает “работу” всеобщего метода случайного поиска в процессе рецепцииуправления на популяциях. При этом, отношения изменений углового положения плоскости закрутки, шага и скорости ее перемещения, отражающие характер рецепции, одновременно находятся во временных отношениях обеих процессов: шаг и скорость перемещения плоскости закрутки характеризует время эволюции, а ее угловое положение – время коррекции. Характер отношений во времени между спиралями эволюции и ее коррекции, оказался сходным (но не более!) с отношением масштабности, фрактальности физического пространства. Удалось численно оценить некоторое его подобие - календарный ““генетический фрактал“, находящийся в отношении биологического “собственного” времени коррекции и “реального”, ньютоновского времени эволюции. Эта величина, оказавшись постоянной, совпала с основанием десятичной системы счисления. При этом, однако, осталось не выясненным – является ли такая “генетическая фрактальность” корректируемого процесса следствием внеземного вмешательства в эволюцию, или собственным свойством организма, или это проявление отношения временной синхронизации, направленного на согласованность достижения множества целей коррекции множеством исполнительных геномов. Мы склонны полагать последнее, ибо появление “производственной”, и при этом нерегулярной спирали коррекции, связано с непрерывной временной синхронизацией множества процессов рецепции на ее витках. Результат этой синхронизации отображается пространственной картиной совмещения конечных условий каждого выполненного витка коррекции с начальными условиями каждого ее последующего витка. Это отражает непрерывность рецепции на популяциях.
“Календарная” дата первого посещения ВВ нашей планеты, отразилась моделью своеобразно - как цикличная с кратностью равной степени фрактала, и с точностью до коэффициента кратности 1,2,...,6 (1x10, 102x10,... 103x10,...106x10 лет) совпадающей с датой, сообщенной визитерами. Ограничение коэффициента кратности значением 6, объективно определяется архитектурой модели, а относительно объекта рецепции оно проявляется как календарное ограничение, выводящее вероятную дату первого посещения за пределы логичности: кратность фрактала равная 7, отрицает реализацию рецепции до появления клонируемого прародителя. Отношения физической возможности отразились через отношения модели.
Полученный результат означает, что смоделированные даты посещений цикличны и определяются кратностью степени фрактала постоянной величины и ни одно из значений кратности не имеет предпочтений. Поэтому, с равной степенью вероятности можно говорить о первом посещении Земли, например 10000 лет тому назад, как заявили ретикуляне или 10000000 лет тому назад, относя его ко времени появления Kenyapithecus или ко времени существования Ramapithecus, как считают дарвинисты, прародителя человека. Важно, что календарные сведения о посещениях в качестве исходных данных в модели, ни в каком виде не закладывались и получились как отраженные собственными отношениями граф-модели, хотя и требующими интерпретации.
Причина неопределенности модельного результата, связанной с отношением кратности временного фрактала, отраженным в модели, осталась не выясненной. Можно было бы для простоты предположить, что она связана с особенностями календаря планеты Ретикули-4, по которому исчисляли время посещений визитеры, оказавшимися “вложенными” в управляющую информацию рецепции. Однако ученые утверждают, что ими выявлены новые фундаментальные свойства того, что мы называем “временем”, и они оказываются способными, если и не объяснить, то интерпретировать этот феномен.
Так, ученые Дж.Гибс и физик Х.Хармут утверждают, что для описания физических явлений использование ньютоновской модели регулярного времени не является необходимым, ибо время “порождается” событиями и является относительно событий вторичным. В нашей модели отношение кратности фрактала - некоторого свойства субстанции времени (как и самого фрактала), проявилось как вторичное от первичных событий рецепции. События рецепции, гипотетически изменив воспринимающую систему – геном ДНК, привели к его необратимому устойчивому изменению, реализующему заданное управление объектом вмешательства. Ньютоновская система отсчета времени в феномене управляемой эволюции, при этом отразилась как вторичное в “самостоятельную” – циклическую, на спирали эволюции, систему отсчета времени коррекции.
Фрактал - суть отношение не времен эволюции и коррекции, а систем их отсчета, отражающее характер отношений цикличности на популяциях в их собственных системах отсчета времени. Но отношения на системах отсчета времен отразилось через кратность фрактала и в отношения цикличности интервалов времен, отсчитываемых с начала каждого из витков коррекции. Это хорошо согласуется с картиной календарных следований циклов эволюции и витков ее коррекции и подтверждается численными характеристиками их потоков.
Заметим, однако, что такое отражение еще не дает оснований для подтверждения гипотезы цикличности собственно времени, его физических “провалах” или “сжатиях”, о чем любят писать фантасты. Здесь мы встретились с различными системами индикации, отсчета, а система отсчета времени не есть субстанция Время. Тем не менее, это согласует версию Хармута и наше предположение об использовании ВВ системы отсчета “самостоятельного”, первичного времени для указания момента первого посещения Земли. Визитеры посетили Землю, отсчитывая дату первого посещения - 10000 лет тому назад в собственной системе отсчета. Если такая система была действительно использована визитерами, не исключено, что она применяется ими и в их рутинной деятельности.
Возвращаясь к вопросу о коррекции эволюции в целом, заметим, что для ее реализации ВВ должны были задаться стратегиями управления и его коррекции – структурой управляющей информации, вносимой в ДНК (надо полагать, она была им доступна), а также адресами геномов, реализующих эту стратегию, либо иметь средства поиска последних. Не исключено, что случайный поиск, как наиболее естественный, был характерен для всех этапов рецепции.
Именно так можно сегодня, в самом общем виде, представить вероятный механизм коммуникации в системе ДНК-ВВ и его реализацию в целях управления эволюцией в том случае, если геномузыка существует реально и, если, фантастически невероятные 65 регулярных визитов инопланетян, реализующих такую же баснословную 70-ти кратную коррекцию эволюции, и в самом деле, состоялись. Но тут можно повторить вполне резонный и столь же недоуменный вопрос: а если они не состоялись, то что породило совпадение результата моделирования с информацией, заявленной ВВ, если их данные в модель в явном виде не закладывались, а моделирование привело к результату, не только совпадающему с контрольным, но и вскрывающему целую систему породивших его отношений? Могла ли стать система этих отношений известной без модельного эксперимента, т.е. априорно? Мы уже отмечали, что ответ на такие сомнения только один: наше моделирование могло быть предварено либо с целью породить сенсацию и придать ей правдоподобное, верифицируемое содержание, либо с целью распространения столь же правдоподобной дезинформации о посещениях Земли инопланетными визитерами. И хотя наука знает много путей для обоснования (и опровержения) такого рода заявлений, в силу уникальности подхода, примененного нами к анализу информации ВВ, мы все же должны поставить эту версию под сомнение. Видимо, такие совпадения, как и молчание по этому вопросу газеты “Совершенно секретно“ не могут быть случайными. Правда, она позднее опубликовала сведения о зловещей масонской ложе MJ-12.
Мы отличали воздействия управления эволюцией от воздействий ее коррекции, полагая, что при своих посещениях, визитеры осуществляли коррекционные вмешательства по принципу обратной связи от достигнутых изменений вида относительно целевой функции, внесенной пачкой импульсов управления первого посещения. Это означает, что в нашей модели “визитерские” стратегии коррекции моделировали управление стабилизации относительно заданной целевой функции. Мы отличали также операторы естественной эволюции от стратегии внешнего вмешательства и рассматривали, первые - как операторы естественных технологических процессов, подлежащих коррекции, а вторые – искусственных технологий “производственного процесса”, учитывая при этом взаимодействия технологий на структурах моделей.
Терминологическое разграничение, объединяющее эти технологии, введено в 80-х годах академиком А.М.Уголевым. Оно нашло свое противоречивое отражение на моделируемых структурах: отличие живого от искусственного, состоит в природе связей, определяющей управление в живой системе. Уникальность феномена клонирования, независимо от его целей и природы исполнителя, заключается в искусственной регуляции процесса развития живого естественными механизмами. Именно такой феномен отражался моделями, и именно он обсуждается нами. Подобные модели, при соответствующих уточнениях и интерпретациях, могут быть настроены на непосредственное отображение возможных исходов искусственного вмешательства в геном человека.
Полученные результаты, в свете последних сообщений ученых, подтверждают, что передача сигналов, определяющих регуляцию в живой системе извне, могла осуществляться внеземными визитерами путем внесения непосредственно в ДНК информации, представимой цепочкой управляющих операторов в виде последовательности пачек цифровых кодов ДНК, соответствующих геномузыке. Вполне допустимо, что гипотетическое внесение ими стратегий управления именно средствами геномузыки, было для них наиболее простым, если не единственным, путем решения вопроса. Для нас этот путь представляется сегодня практической возможностью организации трансляции извне управляющей генной информации (УГИ) геному, локализованному в той или иной хромосоме ДНК и способному реализовать ее рецепцию. И здесь мы приблизились к заключительной и главной цели нашего повествования: оно направлено ни на доказательство ни на разоблачение заявления визитеров.
Будем под УГИ понимать сообщение, направленное на восприятие управления адресатом-геномом, рецепция которого приводит к необратимому устойчивому состоянию сохранения функции генной системы управлять целевой функцией гомеостазиса – системы поддержания жизнедеятельности, в первую очередь связанной с недопущением ее переключения в программную систему антигомеостазиса – запуска и развития злокачественных опухолей. Это означает, что мы говорим о вмешательстве в процесс развития опухолей на уровне управляющих инициализацией и развитием программ. Мы предполагаем, что, зная систему кодирования ДНК, быть может, и ту, которую нашли японские ученые, возможно воспроизведение “производственных” воздействий рецепции, аналогично тому, как это могли делать при своих визитах ретикуляне. Такая уверенность базируется на знание структуры, состава, характеристик и параметров распределенной клеточной компьютерной системы озлокачествления – антигомеостазиса и ее элементных, сетевых параметров и динамизмов, полученных, методами общей теории систем. Эти знания обуславливают программные и аппаратные возможности достоверного и относительно несложного мониторинга процесса озлокачествления непосредственно по всему множеству клеток, образующих эту биологическую систему и участвующих в процессе озлокачествления.
Численные индикаторные оценки предупредительного момента программного отключения сформировавшейся к запуску системы антигомеостазиса, и ее возвратного переключения на систему гомеостазиса, а значит и прерывания ее функции, обеспечивают прекращение процесса инициации или, в некоторых клинических случаях ранней интенсификации роста опухоли. Такие оценки определяются инструментально. Для реализации механизма переключения в системе антигомеостазиса, может оказаться полезной переключающая функция молекул фениленэтиниленолигомеров. Выявляемые инструментальными методами параметры клеток-онкогенов, облегчают адресацию УГИ, реализующей процесс переключения.
Все параметры биологической компьютерной системы антигомеостазиса выявлены с высокой степенью достоверности по верифицированным клиническим данным, и это создает реальные возможности непосредственной, в том числе профилактической борьбы с опухолями. И этому утверждению способствует еще одно открытие ученых-генетиков.
В августе 2000 года средства массовой информации сообщили, что найдены экспериментальные доказательства существования двух генов, мутации которых провоцируют образование злокачественных опухолей, в частности – молочной железы. Эти гены получили название BRCA1 и BRCA2. Ученые пришли к выводу, что поскольку дефектами этих участков ДНК можно объяснить не более 50% заболеваний наследственными формами рака груди, необходимо продолжать поиск других онкогенов, ответственных за возникновение этой болезни. Специалисты из США, Исландии, Германии, Финляндии и Швеции подтвердили этот вывод и сообщили, что, скорее всего, существует третий по счету аналогичный онкоген, локализованный в тринадцатой (из 24) хромосоме человека. Это определяет конкретную адресацию сигнала управления в режиме прямого доступа средствами коммуникативной системы “ДНК - ОВС”, способного вмешаться как в межклеточный, так и в групповой информационный обмен с целью подавления онкогенов. Но отсюда логически следует:
- если человек научился распознавать код ДНК и выделять из него гены, отражая их нотными кодами, то не означает ли это получение реальной возможности распознания собственно целевой программы озлокачествления - антигомеостазиса, отражаемой в генных или иных кодах и именно на генах, локализованных в злополучно пронумерованной роковым числом 13 хромосоме?
Основные параметры сетей межклеточных и групповых взаимодействий в раковой опухоли – мощности, параметры и маркеры полного множества подмножеств ее элементов, группировки подмножеств и параметры их перетоков, реализуемые системой антигомеостазиса, а также параметры локализации опухоли на множествах ее элементов и на всех фазах состояния определены на мере и описаны.
Установлены также порядки, описаны, маркированы и оценены на введенной мере озлокачествления, множества входов и выходов распределенных, в пространстве опухоли и времени ее развития, группировок клеток-активаторов, связанных сетями групповых взаимодействий по реализуемой ими целевой функции и транслирующих сигналы агрессивного управления ростом. И если сказанное ранее относительно прямого доступа к геному ДНК справедливо, то это означает, что сегодня мы обладаем не только средством прямого адресного доступа к программам целевой функции системы антигомеостазиса, определяющей реализацию процесса озлокачествления, но и влияния на нее.
Есть все основания утверждать, что доступ к образующим управляющие сети антигомеостазиса множествам агрессивных клеток, может быть реализован на системе кодирования генной информации непосредственно по каналам распределенной компьютерной биологической системы множества элементов, локализующих опухоль.
Это определяет эффективное естественное средство вмешательства в функционирование обеих систем сохранения – гомеостазиса и антигомеостазиса и, тем самым, целенаправленного и осознанного управления жизненно важными процессами живого организма.
Яков Гельфандбейн
© 1011494 CIPO



Популярное
Пауки в доме — не случайность. Узнайте, что привлекает этих созданий в ваше жилище и как с этим бороться для создания комфортной обстановки.



Последние материалы

Эл № ФС77-72263 выдано РКН 01.02.2018. Учредитель: В.В. Горшенин. Главный редактор: И.С. Новикова.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), home@pravda.ru Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру»
Copyright © 1999-2025, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.