Пространство и биологическое время ракового гомеостата
 Структура и функции клеточного множества
Структура и функции клеточного множества
Вопросы о смысле пространства и времени возникли еще в далекой древности, однако и сегодня это один из сложнейших вопросов науки. Профессор В.Барашенков, говоря о понятиях пространства и времени, определяет их как "структуру сосуществования и изменения всего материального в мире" - от пространства и времени космических миров до микроскопического пространства и времени внутри элементарной частицы. Интерпретация пространственно-временных взаимоотношений отношений на клеточных множествах требует анализа их структуры и функции.
Глубинные свойства общности природы пространства и времени, с необходимостью проявились и на пространственно-временных отношениях клеточных множеств на модели процесса вековой эволюции системы локального гомеостазиса в среду ракового гомеостата. Временной фрактал, как свойство биологического Времени, отражает, с дробной размерностью, понятие Пространства эволюции:
Напомним, что используемый нами инструмент в виде граф-октавы, отображает цепочку 124578 последовательно-сменяемых событий (ситуаций), находящихся в причинно-следственных связях, отображаемых дугами 1-4,4-2,2-8,8-5,5-7, отражающими порядок порождения ситуаций на его вершинах. Вершины 1,3,6,9 — особые. Они отражают динамизмы специфических событий, интерпретируемые в зависимости от характера ситуаций моделируемого мира.
Мы отмечали, что такой граф имеет фиксированную структуру, а его многообразие обеспечивается разнообразиями множества причинно-следственных отношений на дугах и множествах моделируемых ситуаций. Логика этих отношений обуславливается логикой ситуативного мира вершин октавы. Это позволяет, рассматривая циклические процессы, перейти к рассмотрению последовательности дискрет гипотетической коррекции эволюции системы локального гомеостазиса в систему ракового гомеостата. Мы высказали также предположение, что граф-модель есть наиболее полное отображение самодостаточности клеточного множества, и обратно — идентификация с ее помощью любого ситуационного объекта, отражает его самодостаточность.
Рассмотрим набор последовательностей клеточных перетоков, соответствующих 5,10,20,30,40,50 и 60-му виткам эволюции, отражающий отношения упорядоченности управления. (см. "Система локального гомеостазиса как раковый гомеостат"). Каждая из последовательностей 714285, 142857, 285714, 428571, 571428, 714285, 857142 этого набора, как части макросценария процесса, значением своей головной цифры отражает пошаговый исполнительный механизм самосохранения и указывает на текущий индикатор стартовой вершины (клеточной ситуации) перетока, с которой корректируется текущий цикл эволюции. Кроме того, этот набор отражает упорядоченность круговой смены отношений на графе коррекции, определяя на граф-октаве эволюции, ситуации на множествах, подвергающихся коррекции. Граф коррекции рассматривается существующим в пространстве времени десятивитковой Т-повторяемости, синтезируемом внутрисистемными биологическими механизмами. Некоторые специфические особенности системы отсчета этого времени, проявляются на механизмах коррекции. Процесс эволюции отображается в пространстве времени Тц образования ее циклов, которое мы рассматриваем в качестве "реального времени" в Ньютоновской системе отсчета.
Последовательность, составленная из головных цифр — номеров начальных вершин последовательностей перетоков, образует их изначальный порядок 7124578 на граф-октаве. Этот набор отражает порядок смены последовательностей управления, соответствующей организации транспорта на клеточных ситуациях эволюционирующих множеств, и своим последним знаком 8 указывает на завершение формирования механизма сохранения. Первая пара чисел 71 этой последовательности указывает на инициально-круговое замыкание цикла на граф-октаве, реализуемое "по часовой стрелке". Завершение функционирования механизма упорядочения управления эволюцией, отражается вершиной 9 на 65-м витке 13-го цикла эволюции (табл. на рис.2 указанной выше статьи), а полное завершение процессов коррекции, в том числе и переходных процессов на клеточных множествах, отображается полным набором девяток 70-го витка 14-го цикла эволюции. Рассмотрим особенности 15-го, 55-го и 65-го витков эволюции, выделенных в "исполнительной" таблице рамкой, как специфические.
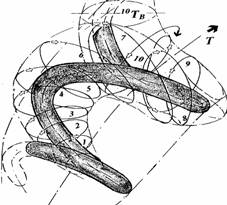
15-й виток, отображаемый последовательностью 214285 ситуаций на вершинах графа, особый. На этом витке отражается первый запуск коррекции эволюционного процесса и единственная за всю эволюцию перемена межмножественных отношений. Эта перемена реверсировала регулярное, структурирующее граф отношение 1-4, заменив его отношением перетока 2-1, упреждающим переход на цепочку 14285, как на цепочку последующих отношений. Это можно понимать, как отображение робастной (грубой) замены первичных на перетоках множеств отношений порождения (1-2), на отношения (2-1) уже на начальном, инициальном этапе эволюции (3-й цикл). Можно полагать, что упреждающее отношение 2-1 отображает "переключатель" отношений управления на организацию генеральной обратной связи [-1(Т)]. Это отношение может задавать ролевую функцию коррекции на ситуации сохраняющего множества. Оно же может исправлять эту функцию на клеточной ситуации, следующей непосредственно за первыми управлениями корректировок на 1-й дискрете 2-го цикла эволюции, результаты реализации которых, не отвечают требуемому качеству самосохранения. Заметим, что такое переключение в состоянии изменить характерразвития, поддерживаемый на ситуациях клеточных множеств механизмом гомеостазиса (возможно существовавшим или, быть может и не существовавшим до 15-го витка эволюции). Вносимая на этом витке ролевая функция самосохранения локального гомеостазиса предназначена для управления механизмом сохранения, антагонистичным механизму гомеостазиса. Такой механизм образует мультиустойчивую систему ракового гомеостата. На титульном рисунке показан характер витков спирали коррекции в трехмерном пространстве времени Т-повторяемости эволюции. Стрелка указывает общее направление эволюции.
Из логики межмножественных отношений следует, что введенные обращением отношения 1-2 переключения, влекут за собой изменение всей последующей системы отношений. Эти изменения происходят не только на клеточных ситуациях (вершинах графа) и перетоках ситуаций (дугах, соединяющих последовательные вершины), но и на структурирующих отношениях порождения — дугах 1-4,4-2,2-8,8-5,5-7 обеих графов — как коррекции, так и эволюции, а также и на отношениях замыкающей дуги 7-1 графа коррекции.
 Измененная на 15-м витке система отношений, фиксирует и определяет цели управления эволюцией сохранения на всех последующих витках коррекции, которые зависят от начальных условий, в свою очередь образуемых соответственно изменяющейся конечной ситуацией на графе каждого витка коррекции. Положение 15-го витка в промежутке 10-20 переходных процессов, отражает срочность запуска переключающего механизма, которая, прерывая переходные процессы предыдущих управлений, игнорирует запрет недопустимости. Длительность дискрет и промежутков в пространстве времени Т-десятивитковой повторяемости, оказались согласованными, а эффективность запущенного механизма корректируется на всех последующих витках эволюции.
Измененная на 15-м витке система отношений, фиксирует и определяет цели управления эволюцией сохранения на всех последующих витках коррекции, которые зависят от начальных условий, в свою очередь образуемых соответственно изменяющейся конечной ситуацией на графе каждого витка коррекции. Положение 15-го витка в промежутке 10-20 переходных процессов, отражает срочность запуска переключающего механизма, которая, прерывая переходные процессы предыдущих управлений, игнорирует запрет недопустимости. Длительность дискрет и промежутков в пространстве времени Т-десятивитковой повторяемости, оказались согласованными, а эффективность запущенного механизма корректируется на всех последующих витках эволюции.
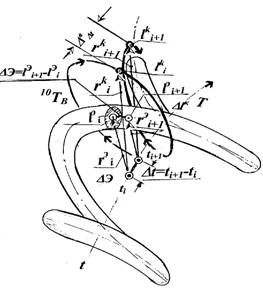
Согласованность масштабов отсчетов времени пространства Т-повторяемости и "реального времени", обеспечивает переменность шага витков и "упаковку" управлений коррекции различной сложности в дискреты одинаковой длительности. Отображение такой согласованности в трехмерном пространстве времени Т-повторяемости для каждой дискреты, представляет собой спиральную траекторию переменного шага (рисунок 1) с витками различной длины, образуемую следом перемещения, от начала отсчета, конца радиуса-вектора также переменной длины. Этот рисунок иллюстрирует (одиночную) дискрету спирали коррекции, навиваемую на спираль эволюции в пространстве времени Т-повторяемости и ее параметры.
 Спираль коррекции образуется следом конца вектора r на каждом шаге i каждого k-го витка в его круговом движении с переменной скоростью "омега" относительно оси эволюции и в поступательном движении с постоянной скоростью вдоль этой оси на интервале времени "дельта" до прибытия в конец витка, совпадающий с началом витка следующего шага i+1. Это создает спиральную "оболочку" из дискрет управления относительно (обладающей двойной кривизной) оси спирали эволюции, элементы которой отражают длительность, в пространстве времени Т-повторяемости, каждого витка коррекции, а ее длина — длительности дискрет управления. Спираль коррекции, внешняя относительно спирали эволюции — причинная, спираль эволюции — порожденная, дочерняя. Она отражает следствие.
Спираль коррекции образуется следом конца вектора r на каждом шаге i каждого k-го витка в его круговом движении с переменной скоростью "омега" относительно оси эволюции и в поступательном движении с постоянной скоростью вдоль этой оси на интервале времени "дельта" до прибытия в конец витка, совпадающий с началом витка следующего шага i+1. Это создает спиральную "оболочку" из дискрет управления относительно (обладающей двойной кривизной) оси спирали эволюции, элементы которой отражают длительность, в пространстве времени Т-повторяемости, каждого витка коррекции, а ее длина — длительности дискрет управления. Спираль коррекции, внешняя относительно спирали эволюции — причинная, спираль эволюции — порожденная, дочерняя. Она отражает следствие.
Путь начала радиуса-вектора r на каждом шаге i k-го витка в его переносном относительно начала спирали эволюции движении, определяет отсчет ti реального времени. Количество витков коррекции, переменная величина шага "дельта" и продолжительности "дельта"t каждого из витков, синхронизируются заданной целевой функцией управления. Путь этого же конца радиуса-вектора, но в его круговом движении, определяет отсчет времени в пространстве Т-повторяемости. Конкатенация (последовательное сцепление) траекторий витков в последовательности реализаций, отражает динамику процесса эволюции на дискретах. Конкатенация траекторий дискрет на витках, отражает эволюционный процесс сохранения. Временному интервалу "дельта"t шага коррекции во времени Т-десятивитковой повторяемости, соответствует шаг эволюции в реальном времени "дельта"t на ее спирали, который является отображением витка коррекции на его шаге.
Путь перемещения начала радиуса-вектора r на каждом шаге i k-го витка в переносном (поступательном) и его конца в круговом (относительном) движениях, задает отношение масштабирования времени пространства Т-повторяемости и "реального" времени и формирует соответствующий механизм текущей синхронизации. Это масштабирование производится на оси эволюции, как на едином носителе временной информации.
На рисунке 2 показаны соотношения на витках коррекции и эволюции. Видно совмещение конечных условий выполненного витка коррекции радиуса-вектора r на i-м шаге k-го (4-го) витка относительно спирали радиуса r-эволюции на этом же шаге, в момент t с начальными условиями i+1 последующего (5-го) витка коррекции, относительно спирали эволюции для последующего момента i+1 времени.
Представление эволюционного процесса с обратными связями в цепи управления, при попытке оценки календарных сведений, определяющих возможное начало эволюционного процесса самосохранения, отобразило существование отношения, имеющего характер масштабности между реальным временем, отсчитываемом в системе Ньютона, в котором отображаются витки эволюции и биологическим временем, в котором отображаются витки ее коррекции, как результата рецепции. Такое отношение возникает при отображении коррекции (причина) в трехмерном Ньютоновском временном пространстве эволюции спиралью (следствие), внешней относительно ее витков и "навитой" на нее (титульный рисунок).
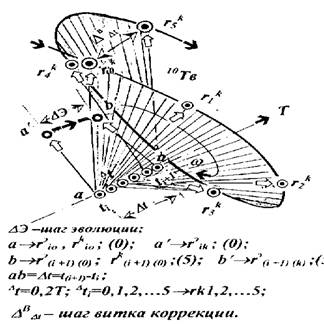
Спираль коррекции характеризуется случайным и неравномерным во времени рецепции шагом закрутки, плоскость которой непрерывно изменяет свое угловое положение в пространстве, одновременно перемещаясь вдоль оси витка эволюции со случайной скоростью шага закрутки. Радиус-вектор спирали, модуль которого также случаен, вращается в переносном движении плоскости закрутки в направлении развития эволюционного процесса. Навитая спираль коррекции в явном виде отражает свое внешнее относительно спирали эволюции происхождение, а также характер "производственного вмешательства" в ее "независимый" от коррекции процесс.
Характерное относительное положение спиралей коррекции и эволюции, присуще специфическим отношениям энергетики пространства и времени, открытым основоположником теории физических свойств времени Н.А.Козыревым. Такие отношения определяют энергетический фактор, как одну из компонент, определяющих характерные признаки самодостаточности сложной системы.
Логично предположить, что случайность значений множества элементов "образующих" геометрии спирали коррекции (линейных, угловых, скоростных, временных параметров перемещения), отражает "работу" всеобщего метода случайного поиска в процессе рецепции стратегии управления на клеточных множествах. При этом, отношения изменений углового положения плоскости закрутки, шага и скорости ее перемещения, отражающие характер рецепции, одновременно находятся во временных отношениях обеих процессов: шаг и скорость перемещения плоскости закрутки характеризует время эволюции, а ее угловое положение — время коррекции. Характер отношений между спиралями эволюции и ее коррекции во времени, оказался сходным с отношением масштабности, фрактальности физического пространства. Удалось численно оценить некоторое его подобие — "календарный генетический фрактал", отражающий отношение биологического "собственного" времени коррекции и "реального", Ньютоновского времени эволюции. Эта величина, оказавшись постоянной, совпала с основанием десятичной системы счисления. При этом, однако, осталось не выясненным — является ли такая "генетическая фрактальность" корректируемого процесса, следствием процесса корректирования эволюции, или отражением ее собственного свойства, или это проявление отношения временной синхронизации, направленного на согласованность достижения множества целей коррекции множеством исполнительных механизмов сохранения. Мы склонны полагать последнее, ибо появление "производственной", и при этом нерегулярной спирали коррекции, связано с непрерывной временной синхронизацией множества процессов рецепции на ее витках. Результат этой синхронизации отображается пространственной картиной совмещения конечных условий каждого выполненного витка коррекции с начальными условиями каждого ее последующего витка, что и отражает непрерывность рецепции на популяциях (рис.2).
"Календарная" дата начала коррекции эволюции отразилась моделью своеобразно — как цикличная, с кратностью равной степени фрактала и с точностью до коэффициента кратности 1,2,...,6 (1x10, 102x10,... 103x10,...106x10 лет). Ограничение коэффициента кратности значением 6, объективно определяется архитектурой модели, а относительно объекта рецепции оно проявляется как календарное ограничение, выводящее вероятную дату начала коррекции за пределы логичности: кратность фрактала равная 7, отрицает реализацию рецепции до появления прародителя Homo Sapiens. Отношения физической возможности отразились через отношения модели.
Полученный результат означает, что смоделированное календарное время коррекции циклично и определяется кратностью степени фрактала постоянной величины и ни одно из значений кратности не имеет предпочтений. Поэтому, с равной степенью вероятности можно говорить о начале формирования системы локального гомеостазиса как ракового гомеостата, например 10000 лет тому назад, или 10000000 лет тому назад, относя его ко времени появления Kenyapithecus или ко времени существования Ramapithecus, как считают дарвинисты, прародителя человека. Заметим, что календарные сведения о таком событии в качестве исходных данных в модели ни в каком виде не закладывались и получились как отраженные собственными свойствами граф-модели, хотя и требующими интерпретации в терминах отношений порождения на клеточных множествах.
Причина неопределенности модельного результата, порожденной отношением кратности временного фрактала, осталась не выясненной. Однако ученые утверждают, что ими найдены новые фундаментальные свойства того, что мы называем "временем", и эти свойства оказываются способными, если и не объяснить, то как то интерпретировать этот феномен.
Так, ученые Дж.Гибс и Х.Хармут утверждают, что для описания физических явлений использование ньютоновской модели времени не является необходимым, ибо время "порождается" событиями и является, относительно этих событий, вторичной категорией. В нашей модели отношение кратности фрактала, как некоторого свойства сущности времени, проявилось как вторичное от событий рецепции (первичное). События рецепции, изменив воспринимающую систему, привели к ее необратимому устойчивому изменению, реализующему заданную коррекцию объекта вмешательства. Ньютоновская система отсчета времени в феномене управляемой эволюции локального гомеостазиса, отразилась при этом как вторичное в "инструментальную" - циклическую на спирали эволюции систему отсчета времени коррекции.
Фрактал или то, что мы назвали "генетической фрактальностью", суть отношение не времени эволюции и коррекции, а лишь согласованности систем их отсчета. Оно отражает характер отношений цикличности на клеточных множествах в их собственных системах отсчета времени. Но отношения на системах отсчета времени отразились через кратность фрактала и в отношения цикличности интервалов времени, отсчитываемых с начала каждого из витков коррекции. Это видно из порядка календарного следования витков коррекции циклов эволюции и подтверждается численными характеристиками их потоков.
Заметим однако, что такое отображение не согласуется с гипотезой цикличности собственно времени, его физических "провалов" или "сжатия": здесь мы встретились с различными системами индикации, отсчета, а система отсчета времени не есть категория "Время". Это некоторое отражение свойства его сущности.
Нет сомнения, что формирование механизмов самосохранения, обращающих систему локального гомеостазиса в ранг ракового гомеостата, может быть реализовано только средствами собственной распределенной компьютерной сети, управляющие рекурсивные воздействия которой, носят характер кодированных сообщений, скорее всего — на уровне молекул ДНК.
Обращаясь к рассматриваемой таблице, мы увидим, что 55-й и 65-й витки (11-и и 13-й циклы) эволюции сохранения также имеют особенности. Цифровые последовательности для этих витков, в последних цифрах отличаются от их фактических значений из-за округления (в сторону развития), результата арифметических операций с семизначными последовательностями 857135 и 9285705 как с числами. С одной стороны можно говорить о том, что логика использованного правила округления отражается в логику обсуждаемого предмета и это не отвергает применимость правила, по крайней мере к рассматриваемой области. Но если рассматривать, полученную для 55-го витка коррекции последовательность, как указание на продолжение циклической перемены сменяемых ситуаций коррекции на вершинах, отображаемых знаком после пятого, то можно говорить об отражении семизначной последовательностью 7857135 разрыва цепочки завершающего отношения 1-4 и о замене его корректирующей цепочкой, задающей уточняющие отношения между ситуациями на клеточных множествах вершин 3-5. Такую особенность цепочки можно понимать как отражение изменений собственно механизма коррекции, быть может, введением "тонкой" корректировки, решающей задачи "доводки" качества коррекции на завершающих ее этапах.
Рассмотрим семизначную цепочку цифр, полученную путем перемножения исходной замкнутой последовательности 1428571 на число витков эволюции сохранения, равное 65 и соответствующее предоконечному 13-му циклу. Получим число 92857115 с головной девяткой. Выделяя из него шестизначную последовательность, получим набор 928571.15. Первые шесть цифр набора образуют индикатор текущего, незавершенного 65-го витка 13-го цикла коррекции, начинающегося с вершины 2 и направленного по цепочке 8571 и далее — по не сформировавшейся "остаточной" цепочке .15.
"Остаточная" цепочка.15 отражает управление, реализующее разрыв упорядочения перспективных отношений на следующем витке коррекции. Такой разрыв устанавливается между клеточными ситуациями, сформированными на его начальной вершине 1 и ситуациями на последующих вершинах графа. Это — управляющая "директива". Она устанавила правило останова процесса эволюции и закрепления достигнутой эволюционной ситуации на вершине 5 непосредственно после реализации отношения 7-1. Такая ситуация фиксируется как конечная, а параметры системы сохранения определяются как достигнутые. Это определяет остаточные знаки рассматриваемой последовательности в качестве фиксаторов достигнутого состояния системы сохранения в раковом гомеостате, 55-й и 65-й витки как "доводочные", а 11-й и последующие циклы — как завершающие циклы конечной 5-й спирали управления эволюцией.
55-й виток коррекции функционально связан с ее 15-м витком. Набор знаков 785714, связанный с отражением разрыва цепочки завершающего отношения 1-4, указывает на возникновение не имевшего места ранее ни на графе коррекции, ни на графе эволюции, отношения между ситуациями 7-8. Это может означать, что исполнительные клеточные множества на ситуации конечной вершины 7 графа эволюции, физически реализовав все множество предшествующих корректирующих воздействий, в том числе и всех предыдущих циклов коррекции, завершает исполнение полного множества соответствий требованиям ролевой функции, внесенного на 15-м витке. Об этом сигнализирует обращение дуги с вершины 7 на вершину 8, ролевая функция которой — завершение готовности создаваемого аппарата выполнять заданную ролевую функцию.
Это одновременно отражает готовность, по завершению переходных процессов эволюции сохранения на ситуациях, соответствующих вершинам 5714 этого графа коррекции, завершить и ее 55-й виток. При этом, на переходных процессах (55-60) витков интервалов недопустимости, завершается информационно-временное согласование механизма уже введенного аппарата сохранения с другими механизмами локального гомеостазиса, обеспечивающими его функционирование в статусе ракового гомеостата. Дискрета 60-го-70-го витков отводится для фиксации и отключения управления.
Таким образом, 55-й виток может характеризоваться, как завершающий задание всех предыдущих управлений и сигнализирующий об этом. Если на 15-м витке экстренно вводились коррекции многообразий и в их числе алгоритмы механизма самосохранения локального гомеостазиса, то на 55-м витке, так же экстренно отменялись дальнейшие управления, с одновременным сохранением управлений, реализованных на предыдущих 50-ти витках коррекции и образовавших 5 спиралей коррекции эволюционного процесса в "реальном" времени. При этом шаг каждой спирали зависит от характера изменения "навивки" витков его коррекции в пространстве времени Т-повторяемости.
Управление процессом сохранения и внутренние сигналы коррекции до 15-го цикла, могли быть простыми реакциями на воздействие внешней среды и не преследовать целей совершенствования системы сохранения. Можно предположить, что регулярность циклов коррекции отражает возможности клеточных механизмов воспринимать управляющие воздействия, независимо от их происхождения. Об этом свидетельствует положение обеих — 15-го и 55-го витков на регулярных интервалах недопустимости. Положение 15-го витка 3-го цикла коррекции связано с характером вмешательства, имеющего цель обзора и учета ретроспективы.
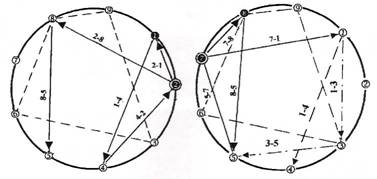 Первый граф на рисунке 3 отражает обращение отношения 1-2, вводящее учет ситуационной ретроспективы, второй — отмену коррекции упреждающим управлением 7-8. Отношение 2-1, отражающее обращение отношения 1-2, на графе коррекции можно интерпретировать как установку на запуск процесса коррекции не с клеточных множеств, достигших развития, отражаемого вершиной 2 графа эволюции, а с множеств вершины 1, находящейся в отношениях перетока 1-2 и которые не успели достигнуть второй спирали развития. Это можно понимать и как стремление охватить генетическим управлением полное множество клеток на транспорте, либо как вмешательство, запоздалое во времени относительно нулевых начальных условий на вершине I. Отношение 7-8 графа коррекции 55-го витка интерпретируется как упреждающее управление, отменяющее коррекцию. Характерное положение 55-го витка отражает срочность вмешательства в процесс эволюции, связанного с текущим контролем достигнутого качества коррекции. Такой контроль может осуществляться, как реализуемый системой самоконтроля качества коррекции, оценивающей экстренность внесения и осуществления текущих управлений.
Первый граф на рисунке 3 отражает обращение отношения 1-2, вводящее учет ситуационной ретроспективы, второй — отмену коррекции упреждающим управлением 7-8. Отношение 2-1, отражающее обращение отношения 1-2, на графе коррекции можно интерпретировать как установку на запуск процесса коррекции не с клеточных множеств, достигших развития, отражаемого вершиной 2 графа эволюции, а с множеств вершины 1, находящейся в отношениях перетока 1-2 и которые не успели достигнуть второй спирали развития. Это можно понимать и как стремление охватить генетическим управлением полное множество клеток на транспорте, либо как вмешательство, запоздалое во времени относительно нулевых начальных условий на вершине I. Отношение 7-8 графа коррекции 55-го витка интерпретируется как упреждающее управление, отменяющее коррекцию. Характерное положение 55-го витка отражает срочность вмешательства в процесс эволюции, связанного с текущим контролем достигнутого качества коррекции. Такой контроль может осуществляться, как реализуемый системой самоконтроля качества коррекции, оценивающей экстренность внесения и осуществления текущих управлений.
15-й и 55-й витки отражают запуск и завершение пятидесятивиткового этапа активного управления качеством эволюции — формирования ракового гомеостата, а также причинно-следственных отношений между ними на всех 50-ти витках коррекции. Полное множество таких отношений включает множество подмножеств отношений управления и их реализации, отображаемых множеством графов эволюции и ее коррекции на всем пятидесяти витковом интервале формирования гомеостата.
Роли 15-го и 55-го витков коррекции осуществляются экстренным введением в процесс эволюции механизмов запуска и отключения управлений развитием многообразий функций на клеточных множествах. Термины "виток" и "цикл" относятся к управляющим воздействиям, термин "спираль" - к эволюционному процессу, как к объекту управления. Витки и циклы коррекции отражают причины, спираль эволюции -следствие. Пять спиралей эволюции переменного шага в текущем времени отображаются пятью вершинами 12457 девятиграммы графа-октавы, находящимися в отношении следования. Каждой спирали эволюции соответствует 10 витков переменного шага ее коррекции.
Взаимное соответствие переменности шага витков коррекции и шага спирали эволюции отображает взаимную зависимость масштабов времени Т-повторяемости и реального времени эволюционного пространства. Текущее, регулируемое качество коррекции отображается вершинами базового треугольника 369 девятиграммы графа-октав ее витков, а достигнутое качество — этими же вершинами конечного графа эволюции. Базовые треугольники всего множества графов находятся в причинно-следственных временных отношениях. Итак:
- 55-й виток коррекции осуществляет "доводку" ее качества путем изменения направлений 1-3-5 межмножественных отношений. Он завершает активное управление коррекцией;
- 65-й виток устанавливает правило останова процесса эволюции и закрепления достигнутой ситуации сохранения. Ситуация на вершине 5 следующего витка фиксируется как конечная, а параметры системы сохранения определяются как достигнутые. Это задает, на завершающих витках 55-60 коррекции, их окончательную направленность, закрепляет достигнутый характер целевой функции, заданной на 15-м витке 3-го цикла коррекции и переключает финальные переходные процессы коррекции на их неуправляемое завершение.
- длительность формирования локального гомеостазиса как ракового гомеостата занимает пять десятивитковых интервалов Т-повторяемости или пятьдесят ее витков (15-й — 65-й), образующих в реальном времени 5 из 7 спиралей эволюции сохранения. Из общего количества 70 витков коррекции, 15 первых (1.5 спирали эволюции) образуют ее некорректируемый или слабо управляемый инициальный этап, а 5 ее незавершенных витков, соответствуют.5 спирали эволюции и отражают асимптотические переходные процессы достижения заданного качества, требуемого для того, чтобы локальный гомеостазис приобрел статус ракового гомеостата.
- витки коррекции формируют вокруг оси спирали эволюции, спиральную "оболочку" переменного шага и сложной конфигурации, образующая которой зависит от характера управления. Витки оболочки и витки спирали существуют в разных пространствах времени, отношения между которыми отражают отношения между шагами и масштабом времени образующих их витков. Витки коррекции, как и витки спирали эволюции могут иметь различную длительность в пространстве реального времени;
- интервал Т-повторяемости играет роль интервала управления. Вершины 369 базовых треугольников девятиграммы графов коррекции, отображаясь в базовый треугольник 369 девятиграммы графа эволюции, отражают множество отношений, структурирующих четырехугольник сохранения 3689 девятиграммы графа ситуационного объекта "эволюция самосохранения".
Использование временной информации о механизмах корректировки эволюционного процесса, связанных с миром сохранения, позволяет обсудить некоторые информационные отношения, порожденные взаимным соответствием пространства времени Т-повторяемости и реального времени корректируемой эволюции, связанные с временным масштабированием ее процессов. Так как граф коррекции эволюционного процесса сохранения многообразий функций не отражает календарного времени вмешательства, особую роль в таком рассмотрении играет безразмерный дискретный временной инвариант (ДВИ) естественно-технологического процесса эволюции.
Раковый гомеостат реализует функцию сохранения специфических отношений на клеточных множествах человеческого организма, обусловленную развитостью межклеточных функциональных коммуникаций и возможностями обработки громадных объемов информации на сетях распределенной клеточной компьютерной системы. Проведенный анализ отражает и подтверждает технологическую и структурную допустимость существования механизма самосохранения.
Граф-октава, как граф "единства", отражает строгую структурную детерминированность неразделимости причинно-следственных и пространственно-временных отношений мира на случайных проявлениях его феноменов. Пространственно-временные и причинно-следственные отношения обуславливают друг друга на множестве отношений порождения, независимо от конкретизации предметной области. Обусловленность пространственно-временных и причинно-следственных отношений отражает энергетическую обусловленность фактора самодостаточности клеточного множества в системе ракового гомеостата.
Изучая причинность в мире клеточных множеств, мы неуклонно приближаемся к новому видению и пониманию Природы. Все живое — сложный и единый организм, эволюционирующий по своим непреложным принципам. Одним из важнейших инструментов его изучения является граф-октава — графическое отражение, геометрический "носитель" глубинной обусловленности структуры и функции как важнейших категорий мироздания, обуславливающих энергию временных трансформаций материального мира.
Яков Гельфандбейн
(c) 1011494 CIPO


Популярное
Как приготовить нежное мясо по-французски? Делимся секретами идеального блюда: выбор мяса, маринад, соус и три потрясающих рецепта.



Последние материалы

Эл № ФС77-72263 выдано РКН 01.02.2018. Учредитель: В.В. Горшенин. Главный редактор: И.С. Новикова.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру»
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.