Вековая эволюция самосохранения
"Истинное знание есть знание причин"
 Точная формулировка проблемы познания, данная Френсисом Беконом, определяет роль и место изучения причинно-следственных отношений мира
Точная формулировка проблемы познания, данная Френсисом Беконом, определяет роль и место изучения причинно-следственных отношений мира
Система локального гомеостазиса реализует множество отношений в этом классе. Как естественно-технологическая, биокомпьютерная система, она эволюционирует одновременно с эволюцией человеческого рода. Проследить механизмы вековой эволюции на классе причинно-следственных отношений можно, представляя ее отношения граф-октавой, аппарат которой применялся ранее для отображения клеточных ситуаций и отношений порождения на них.
Механизмы, рассмотренные в предыдущей статье "Система локального гомеостазиса как раковый гомеостат", позволяют предположить, что адаптивные изменения его системы сохранения функций, носят характер корректировок управлений — параметров операторов на перетоках или взаимодействий операторов, или даже реконструкций — полного изменения одного или нескольких управлений. Такое предположение хорошо согласуется с логикой естественно-технологического процесса развития самой структуры отношений порождения, отражаемых граф-октавой. Однако корректировки операторов или полная замена управлений — процесс, требующий определенных ресурсов и времени, который не может быть завершен в единичном акте или цикле сохранения локального гомеостазиса. Как биологическая система живого организма, этот процесс развивается от одного единичного акта к другому от достигнутого качества, он эволюционирует. Можно предположить, что наиболее лабильным механизмом этого процесса является механизм сохранения функции. Именно эволюция придает системе локального гомеостазиса мультиустойчивость и качества гомеостата.
Нас интересует алгоритмический механизм самосовершенствования процесса сохранения средствами, "встроенными" в систему локального гомеостазиса в качестве инструмента достижения и сохранения им свойств ракового гомеостата. Если фальсифицировать предположение о его существовании не удастся, мы получим возможность оценить естественный аппарат гомеостатирования на уровне его клеточной организации.
Логические основания.
Напомним, что граф-октава отображает цепочку 124578 последовательно-сменяемых событий (ситуаций), находящихся в причинно-следственных связях, отображаемых дугами 1-4, 4-2,2-8,8-5,5-7, отражающими порядок порождения ситуаций на его вершинах. Вершины 1,3,6,9 — особые. Они отражают динамизмы специфических событий, интерпретируемые в зависимости от характера ситуаций моделируемого мира.
Сущность граф-октавы, как утверждает I.B.Popoff в своей книге "The Enneagramma of the Man of Unity" (Samuel Weiser Inc.,1978), заключается в том, что граф отражает единство структуры на множестве причинно-следственных отношений мира во всем его многообразии. Автор иллюстрирует это утверждение многочисленными примерами из различных областей природы и деятельности человека. Так как граф имеет фиксированную структуру, его многообразие обеспечивается разнообразиями множества причинно-следственных отношений на дугах и множествах моделируемых ситуаций. Логика этих отношений обуславливается логикой ситуативного мира вершин октавы. Понимая, под отношением порождения, отношения перехода от малых количественных к скачкообразным качественным изменениям на клеточных ситуациях, придадим им смысл причинно-следственных отношений порождения функции самокоррекции эволюции.
Отображения изменений ситуаций сохранения рассмотрим на логической схеме векового развития, как на объекте, реализующем последовательность корректирующих управлений. Сопоставление отображений отношений порождения и порожденных ситуаций на соответствующих интервалах времени, позволит построить представление об управляемости эволюционного процесса механизмов сохранения.
Анализ логической схемы коррекции сохранения, соответствующей этому графу, показал, что наиболее технологична естественная коррекция с использованием генеральной обратной связи (7-1), либо местной обратной связи (4-2), либо коррекция оператора механизма сохранения 8.
Так как эволюция сохранения в локальном гомеостазисе является лишь одной из компонент эволюции многообразия функций организма, она является и неотделимой частью эволюции вида, находится с ней в отношениях, быть может и определяющих ее. Поэтому ранее рассмотренный логический алгоритм, отражая потоки управляющей информации коррекции, ориентирован на структуру эволюции вида Homo Sapiens. Его отображение в виде граф-октавы, приведено на титульном рисунке. Изображения на отношениях этого графа отражают принадлежность эволюционирующих клеточных множеств сохранения ранее существовавшим видам Homo Sapiens.
Управление на выходе блока 8 (вершины 8 на графе) реализует корректирующие поправки и является функциональным, управляя численными отношениями, существующими во времени как на внутриситуационных перетоках, так и на транспорте клеточных множеств. Эти отношения отображаются в инварианты состояния на рекурсивных отношениях порождения и переноса функциональной преемственности.
Множество вершин граф-октавы эволюции и внутриситуационных перетоков на его вершинах, соответствует введенной корректировке управления, отображает реализацию корректировок на множествах клеток, функционально ориентированных на сохранение. Каждому корректирующему управлению или их набору, может быть поставлен в соответствие либо переток,либо транспортируемое множество, либо их набор на графе эволюции, либо даже направленный граф, отображающий в соответствующем времени отношения, возникающие при реализации корректировок.
Перетоки на ситуациях эволюции описываются операторами, которые в ходе управления эволюцией, могли либо сохраняться с измененными параметрами в классе функционирующих операторов корректируемой составляющей, либо быть введены в ином классе, определяемом целями коррекции. Коррекция с изменением класса оператора, может преследовать цели как развития, так и деградации той или иной функции многообразия на ранних стадиях развития самого оператора. Это означает реконструкцию сохранения или коррекцию одной из его компонент. Следовательно, можно предполагать реструктуризацию многообразия функций в ходе эволюции. Но можно представить также и селективную коррекцию отдельных механизмов сохранения, направленных на развитие избранных многообразий, например — ориентированных на повышение агрессивности клеток баланса эквивалентности, задания сагиттального направления роста или придания отдельным подмножествам клеток функций маркер-трассировщика или интенсификации этих функций. Для реализации коррекции требуются определенные мощности клеточных компьютеров, ресурсы энергии, времени и пространства.
Можно предположить, что коррекция избранных многообразий, реализуемая изменением параметров операторов, требует более ограниченных временных ресурсов, чем коррекция развития или деградации целевой функции, требующая замены класса оператора. Исходя из этого можно считать, что временной интервал со времени начала вековой коррекции эволюции до сегодняшнего дня, мог оказаться достаточным лишь для ограниченных коррекций многообразий, не связанных с изменением класса функционирующих операторов. Такие коррекции могут реализоваться как на внутриситуационных перетоках, так и на транспорте, и их исполнение имеет характер непрерывных, хотя бы и дискретных процессов на клеточных ситуациях. Непрерывные переходные процессы реакции на транспорте множеств, отражаются графами эволюции и их характер отличается от характера управляющих воздействий, носящих во времени скорее всего напрерывно-дискретный или быть может, импульсный характер. Дискретность корректирующих воздействий отражается на циклическом множестве граф-октав коррекции, в качестве носителя причины эволюционных изменений. В соответствующие графы эволюции она отражается как следствие. Механизмы функциональной коррекции эволюции носят причинно-следственый характер.
Исходя из такой концепции, можно полагать, что акт внесения корректирующих воздействий, направленных на создание механизма самосохранения (причина), имеет информативный характер задания программ биологической компьютерной системе и не исключено, что для такой процедуры не требуются большие резервы времени. Реализация внесенных коррекций (следствие), наоборот, требует определенное время для физического завершения переходных процессов и оно может быть достаточно большим, ибо связано с процессами на молекулярном и клеточном уровнях. Из этого следует, что гипотетические возможности самоуправления эволюцией сохранения, как процесса оптимизации, существуют, более того — они достаточно широки. Мы, с целью продолжить наши рассуждения, выскажем следующее (осторожное) предположение:
- если наши представления об управлениях на компьютерных биологических сетях обоснованы, то возможности самоусовершенствования локально гомеостазических управляющих программ свидетельствует и об обратной допустимости — расшифровки программных кодов, с последующей заменой программ самосохранения на программы, разрушающие сложившийся раковый гомеостат и реверсирующие процессы его сохранения.
Аппарат рассмотрения и логика реализации.
 Так как управление эволюционным процессом рекурсивно, мы будем рассматривать его отображение в виде цикличных процессов коррекции на специфицированных ситуациях сетей входа и связывающих эти циклы отношениях. Это означает что, следуя логике граф-октавы коррекции, возникновение цикличности смены ситуационных отношений сохранения должно быть отражено замыканием первого цикла путем введения отношения порождения 7-1 и это отношение порождает первый виток коррекции. Каждый из витков коррекции будет иметь своим отображением девятиграмму соответствующего графа, а начало витка отображается головной цифрой последовательности, специфицирующей порожденную клеточную ситуацию входа (рисунок 1). Порядок вершин по периметру девятиугольника, образует цепь причинно-следственных отношений прямой связи. Дуги графа — связывают вершины в порядке последовательности 142857 введенным отношением порождения. Матрица "6x6" - базис определенности, суть макросценарий процесса. Она образуется в результате последовательного шестикратного циклического замыкания процесса отношением 7-1. Кривые отражают изменения первой разности (ПР), первого (ПО) и второго (ВО) отношений сохранения. Наклонная прямая отражает отношение первых и вторых отношений сохранения к первой числовой последовательности матрицы. Заметим, что циклические графы, динамизмы на них, а также причинно-следственные отношения порождения и сохранения, в книге I.B.Popoff'а не рассматриваются.
Так как управление эволюционным процессом рекурсивно, мы будем рассматривать его отображение в виде цикличных процессов коррекции на специфицированных ситуациях сетей входа и связывающих эти циклы отношениях. Это означает что, следуя логике граф-октавы коррекции, возникновение цикличности смены ситуационных отношений сохранения должно быть отражено замыканием первого цикла путем введения отношения порождения 7-1 и это отношение порождает первый виток коррекции. Каждый из витков коррекции будет иметь своим отображением девятиграмму соответствующего графа, а начало витка отображается головной цифрой последовательности, специфицирующей порожденную клеточную ситуацию входа (рисунок 1). Порядок вершин по периметру девятиугольника, образует цепь причинно-следственных отношений прямой связи. Дуги графа — связывают вершины в порядке последовательности 142857 введенным отношением порождения. Матрица "6x6" - базис определенности, суть макросценарий процесса. Она образуется в результате последовательного шестикратного циклического замыкания процесса отношением 7-1. Кривые отражают изменения первой разности (ПР), первого (ПО) и второго (ВО) отношений сохранения. Наклонная прямая отражает отношение первых и вторых отношений сохранения к первой числовой последовательности матрицы. Заметим, что циклические графы, динамизмы на них, а также причинно-следственные отношения порождения и сохранения, в книге I.B.Popoff'а не рассматриваются.
Рекурсия отражает цикличность как причину порождения эволюционных изменений. Эти изменения, как следствие, образуют необратимый, последовательно корректируемый ситуационный объект эволюции — продукт, порожденный коррекцией. Это позволяет предположить, что спиральный характер, вообще присущий эволюции видов, порождается циклическим характером управлений, реакцией на которые может служить функциональное усовершенствование подмножеств элементов эволюционирующего множества.
Таким образом, рассматривается динамизм ситуационного объекта — механизма самокоррекции многообразия функций клеточного множества. Этот объект отображается в замкнутую числовую последовательность 1428571 причинно-следственных отношений порождения на ситуациях замкнутого графа, которые образуют многовитковый непрерывный процесс управления на специфических клеточных подмножествах сохранения. Каждый виток коррекции, как объект циклического моделирования, формируется во времени для каждого корректирующего воздействия или их набора. При этом, структурная (круговая) упорядоченность 124578 смены клеточных ситуаций на перетоках и в транспорте развивающихся множеств графа эволюции, как и его структура в целом, сохраняются. Меняется лишь интерпретация, соответствующая каждому множеству ситуаций на каждой из вершин и соответственно, структурирующих графы отношений. Каждый новый виток корректирующего управления имеет в качестве начальной ситуации на эволюционирующих множествах ту, которая отображается сменяемой головной цифрой последовательности матрицы 6х6 вершин графа в их очередном наборе.
Смысл ситуаций, отображаемых специфическими вершинами 3 6 9 базового треугольника девятиграммы граф-октавы, конкретизируется межмножественными, на вершинах графа отношениями, зависящими от интерпретации мира коррекции. Интерпретация отношений на ситуациях его вершин и перетоках зависит от отображаемых управлений. Любое множество графов коррекции отражается в единый граф эволюции, количество и последовательность графов коррекции отражает завершенность эволюционного процесса. Упорядоченность перетоков и структурирующих отношений неизменна, интерпретация элементов графа эволюции, как и графа коррекции, зависит от отображаемых отношений порождения, в свою очередь определяемых характером коррекции.
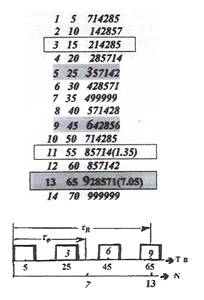 Количество изменений в 5 витков коррекции будем полагать периодом развития, придавая ему смысл цикла. Дальше мы увидим, что интуитивный произвол, допущенный в выборе именно такого количества изменений, образующих цикл, оправдан (как "опирающийся" на число 13 — инвариант механизма локального гомеостазиса).
Количество изменений в 5 витков коррекции будем полагать периодом развития, придавая ему смысл цикла. Дальше мы увидим, что интуитивный произвол, допущенный в выборе именно такого количества изменений, образующих цикл, оправдан (как "опирающийся" на число 13 — инвариант механизма локального гомеостазиса).
Если исходную разомкнутую последовательность цифр 142857, как отражение отдельно взятого витка коррекции, умножать на числа, последовательно кратные договорному числу витков 5 в каждом цикле эволюции (т.е. на 5,10,15,....), получим множество цифровых отображений наборов для каждого цикла изменения ситуаций на вершинах граф-модели эволюции (таблица на рис.2). В отличие от матрицы 6х6 макросценария процесса, эта таблица отражает исполнительный механизм самосохранения на ситуациях вершин граф-октавы. Это — отображение механизма формирования клеточных множеств ракового гомеостата, в свою очередь отраженного в дискретный набор из 10,20,30,40,50,60-го витков коррекции в пространстве Т времени. Так как на клеточных множествах определены соотношения, аналогичные обычным пространственным соотношениям, они образуют обобщенное пространство. Выделим клеточное множество, состоящее из указанного дискретного набора, как пространство "десятивитковой Т-повторяемости" ТП смены последовательностей клеточных ситуаций эволюции.
Циклы 2,4,6,8,10,12,14, начиная с первого, кратные 10 виткам управления, реализуя различное функциональное назначение коррекции самосохранения, отражают ее исполнительный механизм. Т-повторяемость отражает дискретно-непрерывный порядок смены корректируемых ситуаций на клеточных множествах, осуществляющих функцию самосохранения по их циклам за полное время эволюции в пространстве времени десятивитковой коррекции. Исполнение коррекции дискретно-интервальное, также десятивитковое, что отображается последовательностью дискрет коррекции и промежутков переходных процессов на множестве N циклов эволюции самосохранения. Промежутки переходных процессов образуются интервалами между пачками витков коррекции, на которых коррекция предполагается недопустимой. Каждый цикл коррекции отображается своей девятиграммой, которой, в зависимости от характера корректировки, соответствует тот или иной структурный элемент графа эволюции.
В середине каждой дискреты механизма управления, на 5-м, 25-м, 45-м и 65-м витках коррекции, отображающихся в четырнадцати витковую спираль эволюции с шагом, кратным 10 виткам коррекции, проявляется возможность робастного изменения собственно механизма коррекции, путем изменения операторов управления или внесением новых операторов. Об этом сигнализирует "вход" в механизм сохранения на ситуации при вершине 3 девятиграммы граф-октавы (помечен в таблице номером вершины 3), лежащей на основании выпуклого четырехугольника 3689 сохранения второго витка граф-октавы (рис.2). На 45-м витке эволюции со входа на ситуации при вершине 6 той же трапеции, отражается возможность последующей, уже тонкой коррекции этого механизма, путем изменения лишь только параметров ранее внесенных или корректируемых операторов. Это — отражение возможности двухэтапного десятивиткового управления эволюцией, с "доводкой" системы до заданного качества.
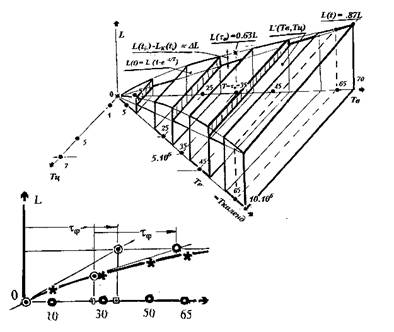 Оценим механизм спиральной Т-повторяемости. Передние фронты дискрет 1,20,40,60-го витков 1,4,8,12-го циклов коррекции — задающие управление. Передние фронты промежутков переходных процессов 10,30,50-й дискреты 2,6,10-го циклов — отсекающие управление и задающие их отработку. 70-й виток 14-го цикла — завершающий. Это определяет положение фронтов внесения десятивитковых (0-10,20-30,40-50,60-70) корректировок и также десятивитковых (10-20,30-40,50-60) промежутков переходных процессов, как отображений реакций на эти корректировки. Завершение коррекции осуществляется на 70-ти ее витках, охватывающих 720 изменений клеточных ситуаций на вершинах графа коррекции и 350 их перетоков.
Оценим механизм спиральной Т-повторяемости. Передние фронты дискрет 1,20,40,60-го витков 1,4,8,12-го циклов коррекции — задающие управление. Передние фронты промежутков переходных процессов 10,30,50-й дискреты 2,6,10-го циклов — отсекающие управление и задающие их отработку. 70-й виток 14-го цикла — завершающий. Это определяет положение фронтов внесения десятивитковых (0-10,20-30,40-50,60-70) корректировок и также десятивитковых (10-20,30-40,50-60) промежутков переходных процессов, как отображений реакций на эти корректировки. Завершение коррекции осуществляется на 70-ти ее витках, охватывающих 720 изменений клеточных ситуаций на вершинах графа коррекции и 350 их перетоков.
Граф корректируемого процесса эволюции, построенный для суммарного текущего числа витков Тв временной коррекции функции сохранения, имея неизменную структуру графа-октавы, отражает 6 его изменений при 5 перетоках на клеточных множествах. Это позволяет отобразить закономерности приращения "дельта"L мощности размножающихся клеточных множеств, обладающих ролевой функцией сохранения, как на всей последовательности смен ситуаций управляемой эволюции, так и на любой составляющей ее части. Такие приращения находятся в причинно-следственном отношении с изменением качества сохранения, внесенным в эволюционный процесс, и могут генерироваться за счет приращения мощности множества клеток сохранения на промежутках переходных процессов.
Полученная зависимость в виде поверхности в системе координат Тв временной коррекции и числа Тц циклов Тв0LТц приведена на рисунке 3. Она отражает приращения L мощности размножающихся клеточных множеств сохранения, "натянутые" на промежутки переходных процессов. Сформированные на этих промежутках множества клеток сохранения пополняют их исполнительное множество на дискретах.
Каждая из дискрет корректировки и соответствующая ей реакция на промежутке переходного процесса, образуют цикл управления синхронизированной длительности, в котором первый полуцикл — внесения корректировок, второй — их реализации (рис.2). Корректировки, вносимые на промежутках переходных процессов — интервалах недопустимости коррекции, прерывая временную непрерывность переходного процесса, носят срочный характер, преследующий цели экстренного корректирующего вмешательства.
Десятивитковая структура спиральной Т-повторяемости циклов коррекции задается дискретным временным инвариантом (ДВИ) процесса эволюции с самоуправлением. Т-повторяемость и дискретный временной инвариант процесса, задающий отсчет внутрисистемного времени, находятся в отношении взаимного соответствия. Отношение ТП -ДВИ отражает пространство дискретного времени десятивитковой Т-повторяемости, синтезируемое аппаратом отсчета времени внутрисистемных биологических механизмов коррекции, как особое подпространство пространства времени Т, в котором совершаются события эволюции.
Положение управлений на 25-м, 45-м и 65-м витках графа коррекции, совпадают соответственно с вершинами 3,6,9 треугольника одноциклической девятиграммы эволюции многообразия функций. Это отражает процесс формирования треугольника механизма самосохранения локального гомеостазиса и определяет 25-й, 45-й и 65-й витки как "ключевые" витки коррекции, реализующие заданное качество сохранения. Вершины 3,6,9 треугольника, образуемого в процессе многоциклической эволюции выпуклым четырехугольником 3,6,8,9 сохранения, отражают клеточные ситуации, "оптимальные" с позиций формирования ракового гомеостата. Совпадение ситуации 60-го витка коррекции в 12-м цикле эволюции, соответственно с 8-й вершиной четырехугольника сохранения в замкнутом цикле (табл.14.1), отражает готовность создаваемого механизма сохранения к функционированию локального гомеостазиса в качестве ракового гомеостата.
Качество коррекции самосохранения в раковом гомеостате отражается как дважды оптимальное, двойным отношением золотого сечения на дугах граф-октавы и близко к асимптотически конечному уровню.
Отношения коррекции ракового гомеостата.
Ключевой проблемой самоуправления эволюцией сохранения локального гомеостазиса, является синхронизация всех процессов коррекции и процедур ее реализации в пространстве времени. Все управления реализуются в пакетах витков. Каждый отдельный виток, как и множества управлений в пакетах витков, в зависимости от характера управления могут иметь различную продолжительность, однако их суммарная продолжительность в пространстве времени Т-повторяемости остается постоянной. Множества витков образуют дискреты из пакетов управлений на витках одинаковой продолжительности в пространстве времени Т-повторяемости. Реализация корректировок, проявляющаяся в цикличности эволюции, приводит к получению ее витков с неравномерным шагом.
Это отражает реализацию некоторого технологического процесса, носящего характер частотной "модуляции-демодуляции" витков коррекции переменной длительности, "упаковываемых" в пакеты равной продолжительности. Для такой "упаковки" требуется синхронизация процессорного биологического времени Т-повторяемости и текущего, "внешнего" относительно механизма коррекции, времени эволюции. Возникает проблема управления межвременными отношениями масштабных преобразований системы отсчета биологического времени (коррекции) и времени реализации циклов эволюционного процесса (следствия). Будем предполагать, что такая операция осуществима на уровне клеточных сетей биологической распределенной вычислительной системы, а употребляя термин "реальное время", будем подразумевать время Тц, в котором отражаются циклы N эволюционного процесса. В частности, оно может отсчитываться в Ньютоновской системе отсчета.
Рассмотрим, врамках рассматриваемой гипотезы, механизм коррекции качества управления. Зависимость, соответствующая изменению мощности сохраняющего клеточного множества, отражаемого в качество сохранения L от числа проведенных корректировок, образует поверхность с сечением в плоскости Tв0L, достаточно близким к экспоненте с постоянной времени, соответствующей 35 виткам эволюции, отображенной на кривой, отражающей изменение мощности клеточного множества сохранения L от числа проведенных корректировок (рисунок 3). Аналогичная зависимость относительно фронтов промежутков переходных процессов (приведена на рисунке), отражает ситуацию линейного роста, которая существовала бы при отсутствии управления коррекции на ее дискретах.
Количество пятивитковых циклов, укладывающееся во время, соответствующее постоянной времени экспоненты и отражающее, тем самым, общее время переходного процесса корректировки эволюции в реальном времени, равно семи (рис.2). Выражение для экспоненты изменения мощности клеточного множества сохранения L от числа проведенных корректировок, записывается в функции числа витков эволюционного процесса. В этом выражении, при t равном постоянной времени, L(t)=0.63L, a при t равном двум постоянным времени и 70 виткам эволюции, L(t)=0.87L. Величина 0.87L определяет максимально достижимое качество сохранения. Постоянная времени экспоненциального роста и изменения качества гомеостатического сохранения равна 35 виткам коррекции. Она отражает свойства реактивности функции сохранения ракового гомеостата на внешние воздействия, в том числе — на воздействия, носящие характер управления. Управление качеством сохранения L носит инерционный характер, соответствующий апериодическому управлению, при котором реакция на импульс коррекции в виде дельта-функции (импульсная переходная функция), описывается затухающей экспонентой с той же постоянной времени.
Таким образом оказалось, что корректировка процесса, максимально допустимая отображающей последовательностью ситуаций на вершинах граф-октавы, ограничивается значением равным 0.87 от теоретически возможного финитного значения качества сохранения L. Из этого следует, что управление корректирует механизм сохранения на реализацию экспоненциального (или близкого к нему), ограниченного роста мощности сохраняющего множества. Интересно, что остаток сохранения отражается часто встречающимся в локальном гомеостазисе числом 13%. В частности, это число отражает относительное количество элементов клеточных множеств сохранения, не приобретающих функции механизма локального гомеостазиса. Приращения объема в пространстве времени, ограниченного поверхностями (линейчатой экспоненциальной — эволюционного процесса с сохранением и наклонной плоской — равномерного роста без сохранения), рис.3, соответствуют, при прочих идеальных условиях, текущему приращению качества корректировок в реальном времени.
Разница между корректируемыми в эволюции параметрами экспоненты и неуправляемым качеством равномерного равновесного роста, слабо зависит от тангенсом угла наклона прямой неуправляемого качества для любого фиксированного момента t эволюции. Она выражается в единицах мощности клеточных множеств сохранения и определяет "сигнал ошибки" управления. Этот сигнал в виде потока единичных импульсов на выходе корректирующей системы, отрабатывается как управляющий. Внесенная коррекция в виде оператора управления характеризуется относительно плоскости L'Тв,Тц. В нашем случае это определяет задание слабо изменяемого ("квазипостоянного") приращения L как константы качества, на всех циклах его корректировки. Последняя коррекция определяется на 65-м витке 13-го цикла.
Граф-октава не отрицает того, что самосовершенствование эволюционного процесса локального гомеостазиса собственной клеточной компьютерной системой, может осуществляться по достаточно простому правилу постоянства приращения качества сохранения "дельта"L=const. Такое управление может задаваться импульсной структурой регулярной коррекции как операторов управления сохранением, так и их параметров на каждом витке коррекции, номер которого кратен 5, за исключением 15-го, 35-го и 55-го витков. Заметим, что это не препятствует генерации срочных сигналов экстренного управления и на этих витках интервалов недопустимости.
35-й виток отличается наличием индикатора 4 (табл. рис.2) и последующей серии указателей 99999. В соответствии с трактовкой смысла вершин граф-октавы, это свидетельствует об индикации и фиксации завершения, с одной стороны, переходных процессов ранних, робастных корректировок на этом витке, с другой — о завершении и фиксации этапа формирования обратной связи в системе коррекции. На это и указывает индикатор 4 вершины граф-модели, специфические особенности которой, в этой связи, рассматривались ранее. По видимому, формирование глобальной обратной связи требует минимальных корректировок в объеме робастных управлений 35-го витка коррекции.
Следовательно, на формирование и фиксацию структуры системы коррекции эволюционного процесса локального гомеостазиса, целью которой является придание ему функции ракового гомеостата, как раз и потребовалось 35 ее витков, которые определяют постоянную времени этой системы. Остальное время (30 витков) коррекции, связывается непосредственно с "доводкой" параметров качества эволюционного процесса тонкими корректировками по линии отношений 4-2,2-8,8-5,5-7 и 7-1 граф-октавы. Такая доводка, однако, производится на предварительно структурированной, и"квазинормально" функционирующей корректирующей системе.
Возможность коррекции после 65-го витка обрывается. Это отображается замыканием четырехугольника 3,6,8,9 сохранения, впервые появившимся на последовательности вершин 65 витка коррекции указателем 9 (рис.2). Такое замыкание можно интерпретировать, как полное исчерпание возможностей совершенствования управляемого объекта при заданном качестве сохранения. Система самосохранения получила качества оптимальной системы, а клеточная компьютерная система локального гомеостазиса — статус ракового гомеостата. Эволюция ракового гомеостата отразилась на уровне клеточной организации, как накопление и самоорганизация долгоживущей клеточной мультиустойчивой системы с модальными параметрами. Заметим, что дальнейшие возможности самокоррекции ракового гомеостата связаны с порождением нового качества самосохранения, и в свою очередь возбуждает повторный множественный цикл развития.
Это вполне соответствует представлениям об эволюции, высказанным в работе О. Эстерле "Новое представление о пространстве и времени в рамках целостной парадигмы" (N-T.Ru [Наука и Техника]), а также концепции "физического времени" Н.А.Козырева, проявившейся на отношениях причины и следствия.
Яков Гельфандбейн
(c) 1011494 CIPO



Популярное
Водители, сверлящие катализатор ради "ускорения", рискуют остаться не только без экологии, но и без двигателя.



Последние материалы

Эл № ФС77-72263 выдано РКН 01.02.2018. Учредитель: В.В. Горшенин. Главный редактор: И.С. Новикова.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), home@pravda.ru Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру»
Copyright © 1999-2025, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.