Биоэнергоинформатика на клеточных множествах
 Кризис теории тонкого мира
Кризис теории тонкого мира
В топологическом пространстве характеристической пирамиды локального гомеостазиса четко выделяются витальные клеточные подмножества древесного роста PROGESSION VITAL, как разделяющие жизнь и смерть, живое и неживое. Концепцию "живое-неживое" связывают с понятием биоэнергоинформатики. В этой концепции, которую в частности, Г.Шипов рассматривает как производную теории "торсионных полей" теоретической физики, понятия энергии и информации тесно связываются с такими психофизическими характеристиками, как сознание и отличие живой субстанции от неживой.
Мы не будем говорить о сознании, тем более как о психофизической характеристике, но памятуя, что какими бы фантастическими не представлялись бы предположения этой, беспощадно критикуемой ныне теории, они требуют проверки и глубокого осмысления на фактическом материале. Поэтому представляет интерес рассмотрение, с позиций этой теории, некоторых отношений еще живой материи, представленной клеточным множеством и уже неживой материи, представленной в динамике озлокачествления тем же множеством, но которое, по своим энергетическим и информационным характеристикам, достигло порога витальности.
Биоэнергоинформатика в теории торсионного мира. Как отмечает К.Гурдин, в работе "Госзаказ на мистику", само по себе явление торсионных полей не несет ничего лженаучного — существование подобных полей предсказано еще Эйнштейном. Проблема лишь в том, говорит он, что ни одной лаборатории мира обнаружить их так и не удалось. Явление торсионных полей обсуждалось достаточно широко и на различных уровнях. Мы, в рамках нашего исследования, и следуя концепции биоэнергоинформатики В.Волченко, рассмотрим, отражение биоэнергоинформативных характеристик клеточных множеств в системе локального гомеостазиса.
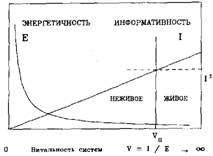
В. Волченко определил условную информационно-энергетическую характеристику живого — "витальность" V через абстракцию, как параметр того, что Г.Шипов определяет "сознанием". Витальность связана с некоторой условной информативностью / живой системы и ее условной энергетичностью Е. Качественное отображение связи этих характеристик, приведено на титульном рисунке, аналитическая зависимость записывается в виде уравнения V = I/E. Согласно его представлениям, для материальных систем существует некоторая "пороговая витальность" Vn, которая разделяет материальные системы на живые и неживые. Отношение между живой и не живой материей рассматривается как отношение переноса энергии и информации.
Под информацией имеется в виду способность к изменению своего собственного состояния или состояния окружающих систем. При этом считается, что изменение состояния может происходить без затраты энергии или с очень малой энергией. Такое представление об информации согласуется с мнением основателя кибернетики Н. Винера, который считал, что информация не является материей или энергией, и что ее способность изменять среду реализует управленческую функцию.
Под энергией подразумевается способность объекта совершать работу. Однако, в концепции В.Волчено не конкретизируется понятие затрат "малой энергии" - собственной, или приобретенной "целевым назначением" от внешнего источника. Именно такое уточнение оказалось необходимым при проведении анализа клеточного роста. Условность понятий энергетичности и информативности, декларируемая биоэнергоинформатикой, достаточно размыта. Эта размытость представляет свободу выбора в согласовании ее атрибутов, в том числе в рамках клеточного мира. Формально понятие "витальности" образуется как производное от отношения характеристик двух клеточных множеств — информативности и энергетичности и, следовательно, от тех условностей, которые обуславливают их понимание. Смысл витальности по В.Волченко — отражение обстоятельств жизненности, она нелинейна, определена на переменных интервалах и стремится в бесконечность.
Биоэнергоинформатика клеточных множеств. Рассмотрим специфику в определениях, касающихся клеточного мира и в первую очередь понятие "условности" информационности и энергетичности. В понятие "условности", с целью сохранения преемственности и создания определенности, мы вкладываем все ранее провозглашенные соглашения и договоренности о рассмотрении численных характеристик на клеточных множествах, как нормированных на собственном максимальном значении, и изменяющихся, следовательно, в диапазоне от нуля до единицы. Это обеспечивает сопоставительный анализ на относительных величинах разного порядка. Считается, что свойство информативности изменять состояние среды, проявляется в технологичности управления клеточным ростом.
Под энергетичностью процесса, мы понимаем затраты условной кинетической энергии, ограниченные по величине, зависящие от объема информации и реализуемые клеточными информационными механизмами. Считается, что они связанны с явлением "подпитки" множеств агрессивных клеток баланса эквивалентности и с механизмами реализации состояния PROGRESSION (апоптоз). Условность такой энергии образуется в терминах энергопотребления по определению, данному в статье "Бигомеостазическое пространство клеточных множеств". Она связана с введением в рассмотрение биологического аналога энергии. В качестве условной массы клеточного множества роста в этом аналоге, рассматривается его мощность на обобщенном показателе озлокачествления (отражающая реальную массу озлокачествленных клеток), а вместо скорости, как характеристики перемещения в физическом пространстве — скорость размножения его элементов, отражающая скорость клеточного транспорта при переходе из состояния в состояние. Такая интерпретация понятия "энергетичности" была сформулирована относительно потребления энергии исполнительными механизмами пересечений фазовых решеток в пространстве состояний.
Применительно к понятию "биоэнергоинформатика" на древесном росте, понятие энергетичности может быть интерпретировано также, в терминах энергоемкости затрат приобретенной энергии на клеточные преобразования и транспорт клеточных множеств в пространстве характеристической пирамиды или полностью определенного дерева роста. Мы рассмотрим основное отношение биоэнергоинформатики как в первой, так и во второй интерпретации энергетичности.
Рассматривая пересечения пространственных фазовых решеток и их ортогональные дополнения, мы получали, на их пересечениях, фазовые траектории по сагиттальным направлениям роста, динамичность которых определяется одновременным кручением и закруткой (двойным кручением) следа перемещения выходной координаты состояния. Можно говорить о создаваемой этим кручением условной инерции роста в ситуации сокращения энергетической подпитки того или иного подмножества на заключительных стадиях процесса, при сохранении резерва клеток, не достигших состояния, стартового для транспортировки на финитный уровень. Такая ситуация отражается при сопоставлении кинетической энергии размножения и клеточного резерва на дереве роста. Свойство кручения — основополагающее свойство торсионного мира, давшее название самой теории. Кручение фазовой траектории изображающей точки, характер его параметров, как и условная инерция клеточного роста — всего лишь отображения физических явлений клеточного мира (как и сама теория торсионного поля лишь отображение мира в сознании человека). Это дает основание полагать, что рассмотрение злокачественного роста с позиций торсионной теории, в силу декларированной условности, по крайней мере не противоречиво провозглашаемым ею принципам.
В.Волченко, используя свободу выбора, по умолчанию рассматривает V-витальность как жизненный фактор. Мы же, при обсуждении древесного роста, рассматривая пространство PROGRESSION, выделили его подпространство PROGRESSION VITAL как некоторое переходное, фиксируя гибельный, роковой характер динамики состояния образующих это пространство клеточных множеств. Потеря жизнеспособности клеток по мере их транспортировки на последующие уровни этого пространства, позволяет определить некоторый порог витальности, за которым следует биологическая гибель клетки. Поэтому, соблюдая преемственность подхода и обеспечивая определенность, понятие "витальность", мы будем понимать именно в этом, а не в ином смысле, и это понимание зафиксируем как L-витальность. Такое осознание L-витальности противоположно смыслу V-витальности: в злокачественном росте L-витальность определяется как обратное понятие, пороговая L-витальность, как условно-договорная величина, разделяет еще живое от уже неживого и имеет характер, существенно отличный от пороговой величины V-витальности. L-витальность ограничена в пространстве роста и времени. Она не образует четкой границы между смежными областями живого и неживого и динамична. В модели L-витальность приобретает наглядность и конкретность — она определяется на мере. В частности, она может быть определена на мере Гомпертца или констант состояния. Витальность и фазовая траектория находятся в отношении однозначного соответствия. Отличие в понимании V- и L-витальности образует основное противоречие в интерпретации формальных отношений биоэнергоинформатики на клеточных множествах.
Структурируя древесный рост в карциногенезе, и исходя из логических условностей и договоренностей, в качестве показателя, отражающего информативность клеточных множеств в динамике роста, мы приняли сложность вершины. Изменение сложности вершины по направлению роста каждой из ветвей, отражает сумму технологий — информационной и исполнительной, и может рассматриваться как накопление условной, в принятом смысле, информации. Однако накопление информации на древесном росте, имеет место не только по направлению роста, но и по его уровням. Характеризуя ветвление, такое накопление, отражает суммы тех же информационной и исполнительной технологий, но вдоль уровня дерева. Исполнительная технология приводит к повышению L-витальности, приближая живую клетку к порогу ее трансформации в неживую, как при переходе ее состояния от уровня к уровню, так и вдоль него. Такой порог, пролегая через все множество клеток, носит, по их множеству, случайный во времени и в пространстве субстрата характер. Понятие четкой грани между живым и неживым размывается.
Сложность вершин, отражая информационно-исполнительные технологии, имеет двоякий смысл. Накапливаясь вдоль ветви по направлению к вершине, она отражает информационность. Накапливаясь вдоль уровня, она отражает левостороннее ветвление кроны. Одновременное накопление — вдоль ветвей и вдоль уровней, отражает L-витальность как редукцию жизнедеятельности. Отражения информативности и энергетичности на дереве роста, аналогичны отношениям ортогональности на пересечениях пространственных фазовых решеток его различных направлений. Если при их пересечении в фазовом пространстве мы получали фазовую траекторию, то используя уравнение В.Волченко, мы связываем информационность и энергоемкость, как затраты энергии, потребляемой от источников подпитки множествами клеток-активаторов баланса эквивалентности и активированными клеточными множествами. L-витальность отражает отношение суммы информационной и исполнительной технологий локального гомеостазиса. V-витальность отражает это отношение в системе гомеостазиса. Отношение L- и V-витальности на клеточных множествах имеет смысл редукции.
Отношения биоэнергоинформатики злокачественного роста. С целью определения отношений L-витальности злокачественного роста и V-витальности гомеостазиса, мы определяли динамичность информативности роста / вдоль каждой ветви дерева и вдоль каждого ее ветвления. Второе отражает информативность разрастания. Сумма сложностей фаз и узлов ветвления на фиксированном уровне, рассматривалась как отражение "текущей" L-витальности, случайной по своему характеру. Последовательное суммирование сложностей узлов и разветвлений ветвей дерева, позволило определить ее нарастание и использовать уравнение В.Волченко для вычисления условной энергоемкости процесса, как величины, обратной энергетичности. Именно такое определение энергоемкости, обеспечило декларированное теорией отражение условно сопредельных пространств живого и неживого.
В соответствии с принятыми ранее условиями, и с целью сохранения определенности, атрибуты роста — информативность, витальность и энергоемкость рассматривались как нормированные на максимальных значениях. В качестве начального уровня во всех случаях принимался узел разрастания PG0 множества эквивалентности. На рис 1 приведены расчетные кривые, точками показаны расчетные значения. Их характер вполне соответствует представлению В.Волченко (титульный рисунок). Однако кривые на этих рисунках, имея достаточно строгое внешнее сходство и совпадая по форме, несут противоположный смысл.
Вначале отметим интересную особенность зависимостей, отражающих условную информативность. Помимо того, что эти зависимости практически линейны (отдельные расчетные значения информативности для различных ветвей 2,3,...7 роста, отличаются не более, чем на одну, и только одна — на две единицы сложности), они конкатенируются в линейную зависимость. Это отражает единый механизм исполнительной технологии и это отражение носит характер информативности роста. Двойное суммирование сложностей вершин дерева по уровням и вдоль уровней, своим нормированным значением, с одной стороны дает практическое совпадение с информативностью роста, с другой характеризует информативность разрастания. Совпадение характеристик информативности роста и разрастания двуедино: оно подтверждает единство управления по всем направлениям активации клеточного поля и единство технологического процесса исполнения.
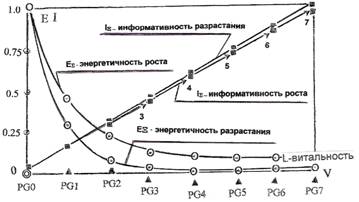 Характеристики энергоемкости древесного роста, вычисленные в своем нормированном отражении, имеют аналогичный характер. Одна из них отражает энергетичность роста, другая — разрастания. Обладая биологически обусловленным максимумом, играющим роль нормирующего значения (0.0769 условных единиц), гарантирующим запуск процесса активации озлокачествления, энергетичность разрастания снижается, асимптотически приближаясь, на уровне PG5 к значению, близкому к 0.0023. Это значение, по договоренности, логично принять за некоторое среднее по клеточному множеству, определяющее пороговую L-витальность (2.4G). Максимум энергетичности роста и его значение для этой пороговой L-витальности, соответственно равны 0.0769 и 0.0092. Таким образом, пороговая L-витальность, при равенстве энергетичностей запуска процессов роста и разрастания, оказалась одной и той же, при четырехкратной экономии энергии разрастания. Это говорит о том, что в смысле энергетических затрат, процесс озлокачествления как в парциальных, так и в общем направлениях роста, параллельно реализуя транспорт клеточных множеств, более энергоемок, чем в направлениях разрастания. С одной стороны, это может обуславливаться необходимостью временной синхронизации транспортных процессов, с другой — спецификой апоптоза, связанной с предпочтительностью подпитки клеточных множеств активации, по сравнению с "пофазовой" подпиткой клеточных множеств агрессии, образующих уровни дерева. Именно такую особенность мы наблюдали на графе агрессии. Не исключено, что, клеточные множества агрессии менее энергоемки, за счет более эффективного использования накопленной инерции роста, в частности — процессов, отраженных в инерцию кручения фазовой траектории.
Характеристики энергоемкости древесного роста, вычисленные в своем нормированном отражении, имеют аналогичный характер. Одна из них отражает энергетичность роста, другая — разрастания. Обладая биологически обусловленным максимумом, играющим роль нормирующего значения (0.0769 условных единиц), гарантирующим запуск процесса активации озлокачествления, энергетичность разрастания снижается, асимптотически приближаясь, на уровне PG5 к значению, близкому к 0.0023. Это значение, по договоренности, логично принять за некоторое среднее по клеточному множеству, определяющее пороговую L-витальность (2.4G). Максимум энергетичности роста и его значение для этой пороговой L-витальности, соответственно равны 0.0769 и 0.0092. Таким образом, пороговая L-витальность, при равенстве энергетичностей запуска процессов роста и разрастания, оказалась одной и той же, при четырехкратной экономии энергии разрастания. Это говорит о том, что в смысле энергетических затрат, процесс озлокачествления как в парциальных, так и в общем направлениях роста, параллельно реализуя транспорт клеточных множеств, более энергоемок, чем в направлениях разрастания. С одной стороны, это может обуславливаться необходимостью временной синхронизации транспортных процессов, с другой — спецификой апоптоза, связанной с предпочтительностью подпитки клеточных множеств активации, по сравнению с "пофазовой" подпиткой клеточных множеств агрессии, образующих уровни дерева. Именно такую особенность мы наблюдали на графе агрессии. Не исключено, что, клеточные множества агрессии менее энергоемки, за счет более эффективного использования накопленной инерции роста, в частности — процессов, отраженных в инерцию кручения фазовой траектории.
 Отношение условных энергоемкостей разрастания и роста, характеризуется "клеточным" энергетическим КПД. На рисунке 2 показано отношение энергоемкостей разрастания и роста. Кривая имеет минимум, положение которого совпадает с положением уровня PG2 древесного роста, на котором формируется разветвление разрастания, образованное множеством клеток баланса эквивалентности. Это отражает повышение расхода энергии разрастания, связанное с активацией клеточных множеств, образующих ветвления кроны древесного роста. Далее, до условного порога витальности PG5, наблюдается линейный рост энергетической эффективности, связанный с использованием накопленной инерции древесного роста. Энергоемкость разрастания, как общая энергоемкость потребления механизмами роста и ветвления, обеспечивает динамику витальности, как глобальности исхода, с опережением относительно парциальной энергоемкости роста. Такое опережение, несмотря на отмеченную энергетическую экономичность клеточного транспорта на фазах ветвления, подтверждает ранее высказывавшуюся мысль относительно лидирующей роли разрастания в пространстве PROGRESION VITAL. Выбор, в качестве порогового значения L-витальности уровня PG5 (2.4G), определен появлением именно на этом уровне, первого множества клеток, характеризующихся значением 0.8 раковой константы. Информативность роста и разрастания, соответствующая такому значению пороговой L-витальности, достаточно велика.
Отношение условных энергоемкостей разрастания и роста, характеризуется "клеточным" энергетическим КПД. На рисунке 2 показано отношение энергоемкостей разрастания и роста. Кривая имеет минимум, положение которого совпадает с положением уровня PG2 древесного роста, на котором формируется разветвление разрастания, образованное множеством клеток баланса эквивалентности. Это отражает повышение расхода энергии разрастания, связанное с активацией клеточных множеств, образующих ветвления кроны древесного роста. Далее, до условного порога витальности PG5, наблюдается линейный рост энергетической эффективности, связанный с использованием накопленной инерции древесного роста. Энергоемкость разрастания, как общая энергоемкость потребления механизмами роста и ветвления, обеспечивает динамику витальности, как глобальности исхода, с опережением относительно парциальной энергоемкости роста. Такое опережение, несмотря на отмеченную энергетическую экономичность клеточного транспорта на фазах ветвления, подтверждает ранее высказывавшуюся мысль относительно лидирующей роли разрастания в пространстве PROGRESION VITAL. Выбор, в качестве порогового значения L-витальности уровня PG5 (2.4G), определен появлением именно на этом уровне, первого множества клеток, характеризующихся значением 0.8 раковой константы. Информативность роста и разрастания, соответствующая такому значению пороговой L-витальности, достаточно велика.
В отличие от концепции В.Волченко, зависимость L-витальности от информативности и условной энергоемкости, привела к "обращению" пространств "живого" и "неживого" и отсутствию четкой границы между ними. Такое обращение есть результат принятых договоренностей, заложенных в построения, в частности — различия в понимании естественной L-витальности на клеточных множествах, относительно абстрактной V-витальности, принятой в биоэнергоинформатике и потребительского характера энергетичности в локальном гомеостазисе. Расчеты показали, что отказ или изменение любой из условностей, сформулированных применительно к системе локального гомеостазиса, приводит к получению закономерностей, совместимость которых, не образует сопредельных пространств живого и неживого, а следовательно, и не может отразить декларируемые непосредственные переходы на них, даже если они и существуют.
Несмотря на такие различия в условностях атрибутов биоэнергоинформатики, в построении частично отразились особенности живых систем, декларированные теорией торсионных полей. Энергоемкость злокачественного роста оказалась существенно ограниченной энергией потребления при подпитке, внешней относительно озлокачествляющихся клеток и при достаточно высокой информативности. В нашем случае она достигает значения 0.75 от своего максимального значения, при условной пороговой L-витальности PG5 (2.4G). Это позволяет полагать, что существование финитных озлокачествленных клеток, в том числе и голых ядер, обеспечивается за счет инерции роста, отображенной закруткой изображающей точки фазовой траектории при ее приближении к фокусу. Энергоемкость клеточного множества, утерявшего жизненные функции, составляет 3-4% от максимального значения и менее, отражая энергию потребления переходных процессов, обуславливающих по Г.Шипову, исчезновение "грубой материальной оболочки — человеческого тела", другими словами — возникновение отношений "тонкого мира". Но в соответствии с теорией тонкого мира, пространство живого возникает из пространства неживого при бесконечном увеличении V-витальности, а не при ограниченном значении L-витальности, интервал развития которой составляет примерно 2.4G, оставляя на динамику переходных процессов к неживому, около 0.9G.
Таким образом, полученное обращение пространств живого и неживого, противоречит принципиальному утверждению теории торсионных полей о том, что живые системы "...обладающие очень высокой удельной информативностью и ничтожно малой удельной энергией могут переходить в тонкоматериальную (или духовную) область жизни, при которой грубая материальная оболочка — человеческое тело, отсутствует".
Такое противоречие ставит под сомнение теорию "тонкого мира" и опровергает ее собственное утверждение, по крайней мере относительно процессов злокачественного роста: L-витальность, в естественном ее понимании противостояния двух антагонистических систем сохранения, летальный исход которого предвосхищен всегда, не может являться формализованной характеристикой духовности, провозглашенной этой теорией. Следовательно, она не позволяет осознать и то, что авторы теории называют иерархией духовных сущностей в эзотерической, предназначенной исключительно для посвященных, и понятной только им, картине мира. Различия в понимании L- и V-витальности и образуют порог "непонимания", разделяющий человечество на "посвященных" и "непосвященных".
Тем более, мертвая клетка не образует субстанцию "душа". Если уже и говорить об этом понятии, то более уместно понимать под этим, некоторое "сгущение" селектированной информативности, реализуемое в памяти распределенной клеточной компьютерной сети, и выводимое из этой — оперативной и долговременной памяти раздельно, при полном завершении постлетальной энергетической подпитки. Импровизируя, можно полагать, что интервал хранения такой оперативной информации составляет около 0.9G от значения L-пороговой витальности.
Логика желания в биоэнергоинформатике. Возникает вопрос: в чем источники указанного противоречия, если картина, соответствующая решению основного уравнения теории, не может быть получена при договоренностях, отличных от принятых? Для ответа на этот вопрос примем, в качестве атрибута энергообеспечения, уже не условную энергоемкость, а условную кинетическую энергию размножения клеточных множеств, и будем вычислять уже не L-витальность по древесному росту, а V-витальность по В.Волченко как отношение равенства между множествами V и I/Е.
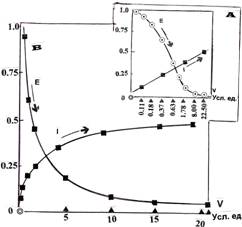 На рисунке (3А) приведены атрибуты V, I и Е биоэнергоинформативного пространства для рассматриваемого случая. В качестве информативности принята ранее определенная зависимость, как средняя между ростом и разрастанием, а в качестве атрибута энергетичности используется условная кинетическая энергия размножения. V-витальность, в отличие от L-витальности, отразилась на неравных промежутках, нелинейно зависит от энергетичности и устремлена в бесконечность. Полученная исходная картина внешне полностью согласуется с положениями теории, но эта, чисто внешняя согласованность, требует дополнительного анализа. Она не подтверждается, как и ранее, декларированным в ней смыслом: пространства живого и неживого обращены.
На рисунке (3А) приведены атрибуты V, I и Е биоэнергоинформативного пространства для рассматриваемого случая. В качестве информативности принята ранее определенная зависимость, как средняя между ростом и разрастанием, а в качестве атрибута энергетичности используется условная кинетическая энергия размножения. V-витальность, в отличие от L-витальности, отразилась на неравных промежутках, нелинейно зависит от энергетичности и устремлена в бесконечность. Полученная исходная картина внешне полностью согласуется с положениями теории, но эта, чисто внешняя согласованность, требует дополнительного анализа. Она не подтверждается, как и ранее, декларированным в ней смыслом: пространства живого и неживого обращены.
На этом же рисунке приведены расчетные зависимости (3В) V-витальности от информативности и энергетичности, как кинетической энергии размножения клеточных множеств. Видно, что при снижении энергетичности, витальность асимптотически устремляется в бесконечность, достигая условной пороговой величины при значении, близком к 22.5 условных единиц. Такой рост V-витальности требует повышения информативности, что также отражается ее характером, как функции V-витальности. Это означает, что повышение жизнеспособности, отраженной в V-витальность, требует снижения энергии подпитки. Это также хорошо согласуется с практикой (явление апоптоза): в системе локального гомеостазиса, как в реальной системе, повышение L-витальности, приближающее ее порог, наоборот, требует повышения подпитки, осуществляемой в субстрате прорастанием кровеносных сосудов в окрестностях активных клеточных множеств.
Отношения биоэнергоинформатики на клеточных множествах, построенные на атрибуте V-витальности, как и ранее, отражают "обращение" сопредельных пространств живого и неживого, присущее реальной картине локального гомеостазиса. Из этого логически следует, что отношения между понятиями V- и L-витальностей распространяются и на соответствующие отношения миров. Неживое следует из живого, живое не порождается неживым, по крайней мере в рамках рассматриваемого подхода. Отношение следования, провозглашенное теорией торсионного мира как желаемое, не нашло своего подтверждения на ярчайшем проявлении связи живого и неживого в процессе перерождения клеточных множеств.
"Торсионный парадокс" тонкого мира биоэнергоинформатики. Сложилась парадоксальная ситуация. Мы применили основное уравнение биоэнергоинформатики, устанавливающее связь между атрибутами, свойственными живой системе, к клеточному миру, в котором реализуется противостояние систем сохранения жизнедеятельности и антагонистической ей системы сохранения разрушения жизнедеятельности. Система гомеостазиса этого мира потеряла устойчивость, в то время, как система разрушения жизнедеятельности — локальный гомеостазис, приобрел свойства системы оптимального управления злокачественным ростом с безграничной устойчивостью. В процессе метаморфозы, управляемой этой системой, пороговая L-витальность определяется естественным путем как его вполне уместный атрибут, устанавливающий грань, хотя бы и условную, между живым и неживым. Динамика процесса отображается информативностью и L-витальностью, как отражением состояния в явном виде и в численной форме на древесном росте. Это позволило, с использованием основного уравнения биоэнергоинформатики, вычислить условную энергетичность (энергоемкость) Е, как функцию условной информативности и L-витальности.
Поиск формального решения через абстракцию, позволяет познать содержание феномена путем выявления элементов его формы. Формальное решение, соответствующее утверждениям теории В.Волченко относительно перехода от живого к неживому в локальном гомеостазисе, соответствует логике злокачественного роста только по отдельным элементам формы, отражая подмену действительного желаемым. Форма, как внутренняя структура, связь и способ взаимодействия частей феномена "живое-неживое" на клеточных множествах, построенная на соотношениях биоэнергоинформатики, оказалась не соответствующей содержанию и смыслу — основе явления и противоречит ей.
Основное расхождение отражается в том, что смежные области еще живого, трансформирующегося в неживое, и область уже неживого, переместились, они поменялись местами. Область живого, в процессе трансформации клеточного субстрата, охватывает пространство, заключенное в границах от начального нулевого значения витальности до ее договорной L-пороговой величины. Такая ситуация, хотя и условно, но адекватно отражает клеточный мир антагонистического противостояния систем гомеостазиса и его результат, но отрицает основные трактовки биоэнергоинформатики: прирост пороговой величины L-витальности не может вести к порождению живого. В локальном гомеостазисе, как в естественно-технологическом процессе, механизмы транспорта непрерывно замещают элементы живого элементами неживого и такое замещение, образуя размытую границу, реализуется, начиная с момента активации. Четкая граница устанавливается лишь в результате гибели последнего элемента клеточного множества и декларирование в качестве такой границы условного "порогового" значения витальности не имеет логического смысла.
Парадоксальность ситуации состоит в том, что применение теоретических положений на реальных данных ярчайшего проявления противостояния живого и неживого — процесса перерождения клеточных множеств, привело к результату, обратному провозглашаемому теорией. Отношение следования биоэнергоинформатики, провозглашенное в качестве одного из основных отношений: "торсионного мира", не нашло своего подтверждения на естественно-технологических клеточных процессах. Это — "торсионный" парадокс: уравнение В.Волченко формально подтверждается, а суждения на полученных результатах, расходятся со здравым смыслом.
Такая парадоксальность, скорее всего, есть следствие того, что при построении теории биоэнергоинформатики, еще не в полной мере были уяснены ее фундаментальные закономерности и логические основания, связанные с провозглашенной размытостью условностей, допускаемых в трактовке ее основных атрибутов, в частности — энергетичности и витальности. Тем не менее, нельзя утверждать и то, что в ее проявлении не сыграл роль естественно-потребительский, затратный характер энергетичности в локальном гомеостазисе. А это — непреодолимое препятствие для использования этой теории в задаче анализа естественно-технологических процессов метаморфозы живого в неживое, при любых, самых реальных условностях.
В.Волченко определил понятие "витальности", в котором свойства множеств определяются через (осторожно) установленное (приблизительное) равенство отношений между множествами. Это — определение через абстракцию при отсутствии четко декларированных условностей. Но определение требует также не только четкого отображения присущих явлению признаков, но и отрицания противоположных признаков для данного явления Только это приводит к определенности — такому качеству сознания, которое свидетельствует о том, что в рассуждение вкладывается одно и то же точное, четкое содержание. Если определенность отсутствует, то свобода выбора условностей приводит к логике желаний, а логика желаний к подмене понятий.
К сожалению, в определении термина "витальность", используемого теорией торсионных полей, нет логического рассуждения, в процессе которого раскрывается содержание этого понятия. Это — следствие отсутствия целостной совокупности суждений о нем, ядром которых являются суждения о его существенных признаках. Обсуждение понятия "L-витальность" дает его понимание через противоположность. Такое понимание позволило внести конкретность не допускающую сомнений, которая, в отличие от условности V-витальности, вносит четкое отображение присущие понятию "витальность" признаков и дает им численную оценку. Это заменило логику желаний, в определении основного отношения "живое-неживое", логикой опровержения, как доказательством несостоятельности базового понятия "тонкого мира".
Парадокс биоэнергоинформативности — парадокс понимания, а следовательно и определения. Парадокс определения — кризис состоятельности теории.
Яков Гельфандбейн
(c) 1011494 CIPO



Популярное
Юрий Кисилёв, известный как певец Юркисс, объяснил, какой должна быть идеальная девушка в его представлении.



Последние материалы

Эл № ФС77-72263 выдано РКН 01.02.2018. Учредитель: В.В. Горшенин. Главный редактор: И.С. Новикова.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру»
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.