Общая архитектура метаморфозы биологического субстрата
Для биоматематиков.
 Механизмы клеточного транспорта и его особые точки
Механизмы клеточного транспорта и его особые точки
Введение аддитивной меры озлокачествления и единицы измерения на ней, определяет четкую интерпретацию всех элементов структуры пересечений пространственных фазовых решеток, как каркаса модели метаморфозы биологического субстрата. Основу такого каркаса формируют сферы шаров гомеостазиса и локального гомеостазиса. Шары соединяются внутренней вертикально-линейчатой поверхностью перетока сложной формы, которая порождается шаром гомеостазиса и рассекает шар локального гомеостазиса. Поверхность перетока делит, покрываемое оболочкой, образуемой пересекающимися внешними горизонтально-линейчатыми поверхностями междушаровое пространство, на два подпространства. Каждое из этих подпространств опирается нижней гранью на общее основание, а их боковые грани образуются плоскостями системы координат, с центром в центре шара единичного радиуса.
Динамизмы процесса отражаются перетоками клеточных множеств из пространств (множеств), покрываемых шаром гомеостазиса в пространство шара локального гомеостазиса вдоль ребра поверхности перетока, а также и через нее — из левого, покрываемого линейчатой поверхностью оболочки междушарового подпространства C, в правое междушаровое подпространство D. Каждый из перетоков отражается, как исполнение функциональной роли целевой функции управления, случайными клеточными множествами отправления случайной мощности, возникает и завершается в случайное время, в связи, или вне связи с другими перетоками. Управление перетоками реализуется на сети межклеточных и групповых коммуникаций. Модель отражает динамику перетоков как тип. Это реализует все основные функции модели.
Начиная с точки касания с шаром локального гомеостазиса, фазовая траектория, как пространственная кривая, обладающая кривизной и получившая стартовый импульс закрутки в момент отрыва от сферы гомеостазиса, на незначительном участке удерживается на его сферической оболочке. Затем, отталкиваясь от нее, она изменяет направление перемещения и направляется к фокусу по кратчайшему пути. Проекция перемещения, текущие значения которого оцифрованы мощностью клеточного множества пересечения в нормированных единицах роста, отражает отношения пересекающихся сагиттальных направлений древесного роста и полностью определяет характеристическую сетку на основании системы координат, как решетку множеств. Пересечения множеств на решетке, определены не обязательно в узлах характеристической сетки.
Кручение траектории перемещения изображающей точки. Кривизна фазовой траектории полностью определяется ее проекциями на основание системы координат фазового пространства (сетку характеристического поля) и ее координатные плоскости, отражая текущие состояния и текущее время процесса. Кручение (вторая кривизна) фазовой траектории — величина, характеризующая отклонение пространственной кривой от ее соприкасающейся плоскости. Кручение считается положительным, если вращение соприкасающейся плоскости при движении вдоль кривой в сторону возрастания параметра от вектора бинормали к вектору главной нормали происходит против часовой стрелки при наблюдении из фиксированной точки Р. Траектория изображающей точки гомеостазиса, обладая кривизной и будучи определенной на поверхности шара, лежит в его диаметральном сечении на плоскости II градации прозрачности ядра и не имеет кручения. Соответственно, главная нормаль (нормаль) к траектории на множестве точек поверхности шара гомеостазиса, проходит через его центр С1.
В момент отрыва изображающей точки от поверхности шара гомеостазиса, траектория получает пусковой импульс "закрутки" и изображающая точка начинает пространственное перемещение. Момент t(0) и импульс закрутки отражают реализацию агрессивных воздействий множества активирующих клеток баланса эквивалентности и образуют первую особую точку траектории. Величина импульса закрутки — функция максимального значения энергии переключения, накопленной переключающим множеством, а текущие затраты энергии на перемещение изображающей точки к фокусу и, соответственно, на закрутку траектории, отражаются биологическим аналогом кинетической энергии так, как это было показано ранее. Есть основания полагать, что отрыв траектории и ее закрутка находятся в отношении порождения с внутриклеточными процессами, отраженными в снижение прозрачности — накопление оптической плотности ядер клеточных множеств.
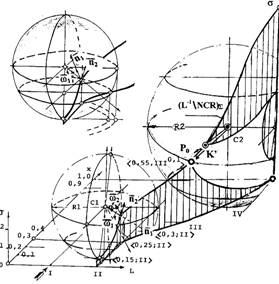 Рисунок 1 иллюстрирует отношения шаров гомеостазиса. Участок P0-K' - зона накопления мощности множеств клеток состояния PROGRESSION, реализующих отрыв траектории от шара локального гомеостазиса и ее закрутку в сторону повышенных значений градаций — снижения прозрачности ядер, по направлению к фокусу. Касание траектории в точке Р0 со сферой локального гомеостазиса, в проекциях на плоскости системы координат xoL, образует точки перегиба. В этих точках, ее проекции из одной полуплоскости, определенной касательной в точке, переходит в другую полуплоскость. Точка касания — вторая особая точка траектории.
Рисунок 1 иллюстрирует отношения шаров гомеостазиса. Участок P0-K' - зона накопления мощности множеств клеток состояния PROGRESSION, реализующих отрыв траектории от шара локального гомеостазиса и ее закрутку в сторону повышенных значений градаций — снижения прозрачности ядер, по направлению к фокусу. Касание траектории в точке Р0 со сферой локального гомеостазиса, в проекциях на плоскости системы координат xoL, образует точки перегиба. В этих точках, ее проекции из одной полуплоскости, определенной касательной в точке, переходит в другую полуплоскость. Точка касания — вторая особая точка траектории.
Кручение фазовой траектории позволяет оценить роль шаров гомеостазиса как сепаратрисных поверхностей. Закручивание траектории для всех пересечений пространственных фазовых решеток порождается в момент t0 начала процесса накопления мощности переключающего множества Q на сфере гомеостазиса.
Рисунок 2 отражает множественный базис пересечений решетки множеств на вероятностной мере, в виде сечения пространственного распределения частостей значений обобщенного параметра N<I,j> озлокачествления и пространства шара локального гомеостазиса, вертикальной линейчатой поверхностью перетока, верхняя грань которой, несет фазовую траекторию.
 Угловые параметры кручения траектории изображающей точки, несут информацию о текущих показателях роста. Точки касания Р0 со сферой локального гомеостазиса и отрыва K' от нее, отражают ключевые динамические факторы взаимодействия фазовой траектории, как пространственной кривой, с сепаратрисными поверхностями архитектуры топологического пространства.
Угловые параметры кручения траектории изображающей точки, несут информацию о текущих показателях роста. Точки касания Р0 со сферой локального гомеостазиса и отрыва K' от нее, отражают ключевые динамические факторы взаимодействия фазовой траектории, как пространственной кривой, с сепаратрисными поверхностями архитектуры топологического пространства.
Накапливая мощность импульса закрутки, переключающее множество клеток Q состояния INITIATION, вследствие внутриклеточных процессов, изменяющих показатели прозрачности ядер, перемещает положение изображающей точки в плоскость II градации L (по направлению, показанному стрелкой на рисунке), и удерживает изображающую точку в этом сечении до момента формирования клеточного множества баланса эквивалентности. Это соответствует моменту отрыва траектории от сферы гомеостазиса в точке с координатами <0.3,0.13, II> и ростом в 0,4G. Отрыв траектории в точке K' от сферы гомеостазиса, с ее одновременной закруткой, отражает границу области разрушения внутриклеточных отношений и реализации гомеостазического переключения. Точка отрыва — третья особая точка фазовой траектории. Четвертая особая точка образуется касанием фазовой траектории изнутри, со сферой единичного шара — фокусом. Ролевая функция особых точек фазовой траектории, как клеточных множеств, — задание межклеточных отношений, определенных целевой функцией и обеспечивающих достижение фокуса по пути наискорейшего спуска..
Изменение пространственной ориентации траектории можно представить также ее изгибанием в двух взаимно перпендикулярных плоскостях, с непрерывным и одновременным перемещением поверхности изменения значений обобщенного показателя озлокачествления — соответственно, в плоскости изменения прозрачности ядер L, и плоскости изменения x ядерно-цитоплазматического отношения.
Закрутка траектории реализуется согласованными взаимодействиями обеих шаров. Вращение вектора главной нормали к траектории, на участке траектории между первой и второй особыми точками, отражает отталкивание траектории изображающей точки от шара гомеостазиса и одновременное притяжение ее к шару локального гомеостазиса, а начиная с третьей особой точки, оно отражает отталкивание траектории шаром локального гомеостазиса в точку абсолютного фокуса. На незначительном участке между второй и третьей особыми точками — касания Р0 и отрыва К', траектория "прилипает" к поверхности шара локального гомеостазиса. Если проявление этого участка не является следствием погрешностей расчетов и построений (точное положение точки отрыва на траектории достоверно установить не удалось), можно предположить, что именно на нем отображается скоротечное создание стартового множества клеток наполнения объема сферы локального гомеостазиса. Это стартовое множество инициирует последующее накопление множеств прибытия клеток на всей трассе изображающей точки от поверхности этой сферы до фокуса, завершающееся раковым взрывом и появлением голых ядер. Об этом, также, свидетельствует картина полностью определенного характеристического поля и характер сечения, образующейся линейчатой поверхностью перетока, многовершинной поверхности частотных распределений, часть вершин которой, оказывается внутри объема шара локального гомеостазиса.
Угол касания соприкасающейся поверхности с траекторией изображающей точки в окрестностях точки Р0 и ее скольжение вдоль поверхности сферы, вплоть до перемены направления вращения вектора главной нормали в сторону фокуса и отрыва траектории, изменяется несущественно и в среднем близок к 55o. Величина этого угла, хорошо коррелируется со значением обобщенного показателя озлокачествления отношения строгого порядка размножающихся множеств, параметром переключающего множества Q состояния INITIATION, а также константой статистической левизны-правизны, рассмотренной ранее. В частности, положению второй особой точки — касания траектории, спроектированной на характеристическую плоскость, соответствует значение ядерно-цитоплазматического отношения, в точности равное 0.55. Это наводит на мысль, что численным значениям угловых параметров фазовой траектории как пространственной кривой, отражающим собственную динамику ее трансформации, соответствует также и нормированная мера, отражающая степень озлокачествления.
Если значения угловых показателей нормировать в пределах от значения угла закрутки проекции траектории в первой особой точке — ее отрыва от шара гомеостазиса, до 900 в четвертой особой точке — достижения абсолютного фокуса, можно увидеть, что угловые параметры собственной динамики фазовой траектории и линейные параметры перемещения изображающей точки вдоль нее (по крайней мере в проекциях особых точек на характеристическую плоскость), находятся в отношении равенства. Это означает, что отношения сходства на клеточных множествах, отражаются в отношения равенства на инвариантах состояния, что и определяет характер проекций, как узлов характеристической сети. Это заключение, однако, нуждается в более тщательной проверке.
Таким образом, феномены кручения и изгиба траектории проявляется как суммарный и совместный результат групповых межклеточных взаимодействий между множествами клеток, покрываемых шаром гомеостазиса и наполняющих пространство шарового сегмента сферы локального гомеостазиса. Эти взаимодействия реализуют движение изображающей точки к фокусу по кратчайшей траектории "наискорейшего спуска". Можно говорить, что отрыв и перемещение изображающей точки возникают как отображение реакции системы локального гомеостазиса (изменений состояний) на систему взаимодействий пересекающихся множеств клеток, создающих клеточную сеть входа Г, а направление этого перемещения осуществляется закручиванием траектории на соответствующей подсети выхода. В рассматриваемом случае такое закручивание задается множеством находящихся в отношении взаимодействий множеств клеток (L-1)сагиттального представления роста, как ортогонального на решетке исходного описания, множеству клеток сагиттального представления роста NCR-ядерно-цитоплазматического отношения. Специфика кручения проявляется по разному для различных пар пересекающихся решеток.
Характер полученных траекторий позволяет утверждать, что различия в групповых межклеточных взаимодействиях, а также собственные динамические изменения сетей выхода, определяющие состав и структуру решеток процесса трансформации, изменяют состав и структуру сетей входа Г в соответствии с процессами транспорта озлокачествляемых клеток. Это отражается в изменении мощности множеств клеток, покрываемых шаровым сегментом шара локального гомеостазиса и это же влечет за собой изменения динамических свойств шаров, как сепаратрисных поверхностей. Характер сепаратрисных поверхностей отражает инициирующую агрессивность множества покрываемых ими клеток баланса эквивалентности, и реализующее эту активность множество клеток баланса толерантности состояния PROGRESSION.
В феномене транспорта, шар локального гомеостазиса, порожденный в момент достижения изображающей точкой положения, соответствующего радиусу сферы в 1.5G (для другой пары пересекающихся решеток — 1.66G), взаимодействием образовавшегося непустого множества трансформирующихся вдоль плоскости перетока и покрываемых его сферой клеток, отображает финитное пространство прибытия. В это пространство транспортируются клетки, межградационная и градационная прозрачность ядра L которых, отлична от значения, определяющего линейчатую плоскость перетока, создающуюся в его динамике. Притягивая изображающую точку, после закрутки до ее касания с поверхностью своей сферы в точке Р0, шар локального гомеостазиса (после непродолжительного удержания), отталкивает ее в направлении к фокусу.
Это отражает два рассмотренных типа динамической агрессивности — "затягивания", свойственного множествам клеток состояния PROMOTION, и "возбуждения" - свойственной механизму подпитки процесса озлокачествления PROGRESSION. Такое, чередующееся воздействие на перемещение изображающей точки, является результатом динамичности вступающих в систему функциональных взаимодействий множеств подмножеств клеток пересекающихся фазовых решеток, обладающих во времени различным типом агрессивности и различными запасами энергии ее реализации.
Не исключено, что подпитка, возникающая как феномен ангиогенеза, отражается (и реализуется) именно на участке "прилипания" P0-K' траектории к поверхности сферы шара локального гомеостазиса. По видимому, она осуществляется не просто в непосредственной близости к узлу [0.6,IV] подпитывающего процесс активации подмножества клеток состояния PROMOTION максимальной мощности 0.86 (точка Е в проекции на характеристическую сетку), но имеет непродолжительный (по изменению состояния субстрата и времени) характер стимула, начиная со второй особой точки Р0 [0.55,III+], на участке Р0-К', порождения этого подмножества.
Сфера шара гомеостазиса и поверхность шарового сегмента шара локального гомеостазиса, образуя сепаратрисные поверхности фазового пространства карциногенеза, отображают систему групповых взаимодействий множеств клеток биологического субстрата, образующих множество его подпространств. Эти взаимодействия первопричинны, и именно они порождают изменения состояний, отображаемых следом перемещения изображающей точки. Взаимодействия сепаратрисных поверхностей и согласованное закручивание фазовой траектории отражает целевую функцию системы локального гомеостазиса как управляющий фактор биологической системы, организующий взаимодействие множеств клеток и направляющий изображающую точку в фокус по кратчайшему пути. Это свидетельствует о наличии, в системе локального гомеостазиса, наряду с признаками многоканальности, также и признаков оптимальности, свойственных системам программного управления. Именно к такому типу систем, можно отнести и систему стабилизации неравновесного состояния — злокачественного роста клеточных множеств.
Двойственная роль притяжения и отталкивания изображающей точки сепаратрисными поверхностями отражает, хотя и дифференцированные в процессе транспорта, но согласованные результаты групповых межклеточных взаимодействий между множествами клеток, покрываемых шаром гомеостазиса и шаровым сегментом шара локального гомеостазиса. Этим отражается согласованность управления. Не исключено, что такое согласование есть проявление оптимальности.
Отображение фазового пространства в топологическое. Фазовое пространство локального гомеостазиса есть некоторый абстрактный объект — результат мысленного вычленения интересующих нас свойств и отношений с позиций общей теории систем. В объеме этого пространства определена, как уже отмечалось, система клеточных подмножеств — открытых множеств, такая, что пустое множество и все пространство являются открытыми множествами, их объединение, и пересечение любых двух открытых множеств также являются открытыми множествами.
Такое утверждение не является вполне строгим. Однако его можно аргументировать тем, что все множество подмножеств клеток — точек, определенных аксиоматическим описанием, содержит точки метрического пространства (например в метриках характеристической сетки или Гомпертца), каждая из которых является внутренней точкой множества, как входящая в него вместе с некоторой своей окрестностью, т.е. рассматривается как множество, содержащее данную точку N<I,j> и открытую окрестность этой точки. Это означает, что все множества исходного описания — открытые. Такое множество может фактически содержать не одну внутреннюю точку, а их совокупность в виде ядра — внутренности множества. Окрестности и внутренность множеств, определенных на узлах сетки характеристического поля, образовались как открытые естественным путем, порожденным робастностью исходного описания показателя прозрачности ядер клеток и рандомизованными методами отбора выборок клинических измерений.
Это означает, что фазовое и топологическое пространства находятся в отношении соответствия: физические (биологические) процессы отображаются в фазовое пространство, которое может рассматриваться как топологическое, элементами которого являются точки. В таком пространстве изучаются такие свойства фигур, которые являются инвариантами, т.е. не изменяются при любых деформациях, производимых без их разрывов и склеиваний — при взаимно однозначных и непрерывных отображениях. Топологическое пространство представляет для нас средство опосредования биологических процессов размножения клеточных множеств на множествах точек, соответствующих множеству образующих его элементов, конкретные параметры которых известны и находятся в отношении упорядочения. Это позволяет уточнить формальное понятие архитектуры фазового пространства карциногенеза как объекта общей теории систем:
- фазовое пространство карциногенеза образуется как топологическое, отображается в объеме, вырезанном пересечением пространственных фазовых решеток из объема единичного шара, внутри которого существуют внешняя и внутренняя сферические поверхности, покрывающие множества клеток гомеостазиса и локального гомеостазиса, играющие роль сепаратрисных поверхностей, соединенных вертикальной линейчатой поверхностью перетока. Поверхность перетока разделяет междушаровое пространство на подпространства клеточного транспорта, образует верхнюю грань пересекающихся множеств, и несет на своем ребре фазовую траекторию, как пространственную кривую — след изображающей точки, собственная динамика которой, характеризуется некоторым множеством структурных инвариантов.
Общая внешняя архитектура такого пространства приведена на титульном рисунке. На нем выделены координатные плоскости отображения текущих состояний (1), текущего времени (2) и вертикальная линейчатая поверхность перетока — транспорта клеток (3). Его общая внутренняя структура определена разделением топологического пространства шарами гомеостазиса — А, локального гомеостазиса — В и секущей плоскостью перетока, — на подпространства C и D. Это разделение отражает характер групповых межклеточных взаимодействий, проявляющихся на пересечениях фазовых пространственных решеток.
Перетоки множеств как клеточный транспорт. Групповые взаимодействия между функциональными подмножествами клеток порождаются в момент t(0) гомеостазического перехода, соответствующего моменту отрыва и закрутки фазовой траектории от сферы гомеостазиса, и существуют, видоизменяясь, в течение всего процесса трансформации. Результат этих взаимодействий проявляется в виде непрерывного перетока и последующего транспорта клеток, из одного подмножества в другое в границах одного подпространства и сформированных стартовых подмножеств — из одного подпространства в иные подпространства, в зависимости от образующихся комбинаций фаз состояний клеток. Непрерывность транспортного потока поддерживается функционированием системы гомеостазиса, продолжающей, в условиях антагонистических отношений с системой локального гомеостазиса, поддержку нарождения множеств нормальных клеток, подвергающихся далее трансформации и пополняющих транспортный "расход".
Топографически, переток множеств клеток, формирующий каноническое состояние, отображается на основании системы координат x0L, как реализуемый по линии, являющейся проекцией ребра разделяющей линейчатой поверхности, несущей фазовую траекторию от поверхности шара гомеостазиса к поверхности шара локального гомеостазиса и далее к фокусу. Мощность перетока характеризуется текущим положением изображающей точки и, соответственно, определяется мощностью подмножеств клеток на пересекающихся фазовых решетках, покрытых интегральным сагиттальным пересечением в объемах подпространств междушарового пространства.
Таким образом, фазовая траектория несет полную информацию о кинетике биологических процессов размножения клеточных множеств. Она характеризуется в топологическом пространстве единицами озлокачествления, выраженными на мере G Гомпертца, а в фазовом пространстве — соответствующими идентификаторами фаз, уровней и страт текущих состояний.
Все, что говорилось выше, касалось картины канонического, завершенного процесса. Однако динамичность текущего процесса трансформации клеточного множества по данным клиники, отраженного в персоналиях, отображается не только характером перетока. Перетоки, непрерывно изменяя отношения между мощностями подмножеств клеток, покрываемых оболочками подмножеств разделяющих поверхностей, изменяют геометрические соотношения архитектуры топологии и, увеличивая объем шарового сектора локального гомеостазиса, одновременно изменяют объемы и конфигурацию междушаровых пространств, отражающиеся в рост протяженности линии перетока.
Линия перетока, определяя фазовую траекторию, отображает также и энергетические отношения транспорта на динамических изменениях мощности множеств клеток, стремящихся к озлокачествлению — достижению фокуса. Построение фазовых траектории по данным клиники, приводит к получению персоналий, отличных от фазовой траектории типа. Сопоставительные процедуры на траекториях типа и персоналий, проводимые на мере Гомпертца, отражают редукцию текущего состояния. Особые точки фазовой траектории, в процессе динамических трансформаций топологического пространства, связанных с развитием процесса, сохраняют инвариантность положения, образуя реперы архитектуры — структурные инварианты.
Не исключено, что на завершающих стадиях процесса может сокращаться и пополнение шара гомеостазиса, покрывающего множество клеток пространства А, за счет снижения мощности множества нарождаемых клеток, связанного с исчерпанием жизненных функций, определяемых нарушением внутриклеточных отношений и активным подавлением механизма гомеостазиса.
Критерии для оценки динамичности перетока, как отображения энергетических отношений, могут быть определены на отношениях в топологическом пространстве, выраженных в гомпертцовской мере. Существует инвариант таких отношений — относительное пространственное положение центров С1 и С2 шаров систем гомеостазиса.
В первой особой точке фазовой траектории отображается акт возникновения взаимодействий между множеством подмножеств клеток гомеостазиса со множеством клеток (L-1)ортогонального дополнения. Эти взаимодействия имеют агрессивный характер типа затягивания, отраженный в узел сцепления <0.3,0.13,II> проекции траектории на характеристическую сеть и отражающий скачкообразность закручивания траектории, как отношение порождения.
Множество голых ядер представлено двумя подмножествами клеток IV градации прозрачности, которая оказалась присущей только таким ядрам. Это отразилось в асимптотическом характере фазовой траектории при ее приближении к фокусу. Особенности распределения частостей ядерно-цитоплазматического отношения при его высоких значениях, связанные с раковым взрывом, обсуждались ранее.
Поверхность перетока, как управляемая полупроницаемая мембрана. Множество клеток подпространства В, покрываемых сферой шарового сегмента локального гомеостазиса, отражает состояние PROGRESSION и кущение дерева роста, образующее на нем область PROGRESSION VITAL. Множество клеток подпространства D отображает состояние PROMOTION ACTIVE, которое в виде ветви этого дерева содержит подмножества клеток подпитки, характеризуется агрессивностью типа затягивания, и совместно с областью PROGRESSION VITAL доопределяет отображение древесного роста.
Это устанавливает отношение соответствия, в динамике текущего процесса трансформации клеточного множества, между фазовыми подпространствами и древесным ростом. Древесный рост рассматривается как управляемая прижизненная форма существования клеточных множеств, при которой первоосновой является ствол, а кущение относительно ствола, рассматривается как персональные рост и разрастание на его ветвях и ветвлениях.
Ствол древесного роста, как его костяк, первооснова, и особые точки, как структурные репера архитектуры фазового пространства — инварианты процесса, находятся в отношении взаимного соответствия.
Поверхность перетока не просто разделяет междушаровое пространство на два динамически изменяемые подпространства: транспорт клеточных множеств через нее, отражает существование особых межклеточных механизмов исполнительной функции транспортных процессов. Исполнительная функция механизма транспорта подобна биологическому механизму осморегуляции и является его аналогом, но действующим на уровне множеств клеток и имеющим специфический характер "макро" осморегуляции. Такая регуляция обеспечивает заданный целевой функцией динамический транспорт клеток через разделяющую поверхность из области С в область D, чем достигается реализация функций активации множеством агрессивных клеток состояния PROMOTION.
Фазовая траектория, как отображение роста текущих мощностей множеств клеток пространства прибытия — линия перетока, отражает функциональные взаимодействия шаров. Можно полагать, что вдоль этой линии, в первой особой точке, стартует функция активации множества клеток баланса эквивалентности состояния PROMOTION, с параллельно активируемым на участке между первой и второй особыми точками, множеством клеток баланса толерантности состояния PROGRESSION.. Такое параллельное подключение активирующих клеточных множеств требует, для отрыва фазовой траектории от поверхности шара локального гомеостазиса, повышенных энергетических расходов, что и обуславливает появление, в окрестностях между второй и третьей особыми точками, энергетической подпитки. Описанный механизм логически следует из рассмотрения полностью определенного характеристического поля (титульный рисунок).
 Разделяющая поверхность перетока может быть определена, как двухсторонняя сепаратрисная поверхность, одна из сторон которой, обладает свойством притяжения, а другая — отталкивания. Двойственное свойство этой поверхности, делает ее функционально подобной полупроницаемой мембране, реализующей односторонний транспорт. Проницаемость поверхности перетока, как полупроницаемой мембраны, может быть оценена на мере озлокачествления, нормированным отношением мощностей стартовых множеств клеток области C, прилегающих в непосредственной близости к ней, к мощности множеств клеток, транспортированных через нее в область D.
Разделяющая поверхность перетока может быть определена, как двухсторонняя сепаратрисная поверхность, одна из сторон которой, обладает свойством притяжения, а другая — отталкивания. Двойственное свойство этой поверхности, делает ее функционально подобной полупроницаемой мембране, реализующей односторонний транспорт. Проницаемость поверхности перетока, как полупроницаемой мембраны, может быть оценена на мере озлокачествления, нормированным отношением мощностей стартовых множеств клеток области C, прилегающих в непосредственной близости к ней, к мощности множеств клеток, транспортированных через нее в область D.
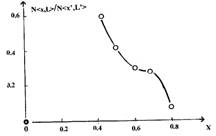
Мембранная проницаемость поверхности перетока, имеет убывающий, S-образно подобный характер. Ее максимальное, начальное значение лежит на некотором удалении извне от сферы гомеостазиса в районе первой особой точки фазовой траектории, и обеспечивается накоплением первичного множества клеток состояния PROgression, порожденного множеством клеток баланса толерантности. Плавно снижаясь, она в непосредственной близости от точки отталкивания шара локального гомеостазиса К', имеет точку перегиба. В этой точке характеристика проницаемости, принимая значение x=0.6, соответствующее значению дисплазионной константы — инварианта процесса, оказывается в положении, находящемся в окрестности, определяющей положение максимума нормированного текущего значения кинетической энергии размножения клеточных множеств. Это позволяет говорить о соответствии энергетических затрат и перетоков, реализуемых в единицу времени, через единицу поверхности перетока, т.е. об их интенсивности. Положение максимума мембранной проницаемости, также хорошо коррелируется с особыми свойствами накопленного значения потери прозрачности ядер в окрестности отрыва фазовой траектории от поверхности шара гомеостазиса, и образованием единицы меры озлокачествления — гомпертца.
Снижение проницаемости "мембраны" перетока, можно толковать, как отображение исчерпания "резерва" клеток, находящихся в левом междушаровом подпространстве C, играющем роль "резервуара резерва". В этом смысле, междушаровое пространство D может рассматриваться как "резервуар расхода". Поверхность перетока играет роль "осмотического насоса" клеточного транспорта. Заметим, что осморегуляция как физический процесс, играет важную роль в физиологических процессах и биологических структурах. Здесь же мы встретились с отображением процесса клеточной проницаемости на модели клеточного транспорта, которое удобно трактовать, в терминах осморегуляции, как ее некоторого аналога.
В терминах теории управления это означает, что множество клеток, транспортированных через плоскость перетока, реализует целевую функцию управляющей системы, отражая функцию "свертки" фазового пространства, играющей роль "множественной" передаточной функции системы управления метаморфозой клеточного множества. Свертку для линейного приближения процессов трансформации клеточных множеств передаточной функцией, мы рассматривали ранее, как "феномен 0.3-0.8" - в задаче анализа вероятностно-динамических преобразований ядерно-цитоплазматического отношения.
В терминах общей теории систем, поверхность перетока определяется как двухсторонняя сепаратрисная поверхность, сторона которой, обращенная в направление ретроспективных значений градации прозрачности ядер обладает свойством притяжения, а в сторону их роста - отталкивания. Верхняя грань этой поверхности отражает интегральный переток, соответствующий множеству состояний и характеризующий множество изменений.
Яков Гельфандбейн
(c) 1011494 CIPO



Популярное
Актёр Максим Матвеев поздравил супругу, актрису Елизавету Боярскую с днём рождения.


Последние материалы

Эл № ФС77-72263 выдано РКН 01.02.2018. Учредитель: В.В. Горшенин. Главный редактор: И.С. Новикова.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), [email protected] Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру»
Copyright © 1999-2024, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.