Форсировки и дегенерация клеточных множеств
 Отображение древесного роста и статус состояния, как меры озлокачествления, позволяют получить отображения второго порядка сагиттальных представлений роста, которые можно определить как функции, отражающие "состязательные" свойства его парциальных составляющих. Это скоростные характеристики процессов. Они отражают их опережающие и форсирующие свойства на отношениях предпочтительности и накопления, как в пространстве состояний, так и во времени, а также накопления значений ядерно-цитоплазматического индекса, отраженного в рост мощности клеточного множества, образующего обобщенный параметр озлокачествления.
Отображение древесного роста и статус состояния, как меры озлокачествления, позволяют получить отображения второго порядка сагиттальных представлений роста, которые можно определить как функции, отражающие "состязательные" свойства его парциальных составляющих. Это скоростные характеристики процессов. Они отражают их опережающие и форсирующие свойства на отношениях предпочтительности и накопления, как в пространстве состояний, так и во времени, а также накопления значений ядерно-цитоплазматического индекса, отраженного в рост мощности клеточного множества, образующего обобщенный параметр озлокачествления.
Ускорение приращения степени поражения биологического субстрата, может отображаться как возрастание скорости активации за единицу времени, полученное в результате сопоставления степени поражения субстрата для различных пар сагиттальных представлений парциальными составляющими направлений роста. Эти отношения образуют параметр, отражающий форсировку или задержку (отставание) процесса в виде приращения его форсировки в пространстве состояний. Приращение форсировки, например, обусловленное парой сагиттальных направлений CanS, отражающего процесс активации методом «прямого» возбуждения и glS – глобального направления, определяют рост и поддержание неравновесного состояния биологического субстрата на 1÷5-м уровнях озлокачествления. Приращения форсировок между всеми парами сагиттальных направлений имеют аналогичный качественный, но количественно различный характер. Это отражает единство управления, хотя и с различной эффективностью, на всем множестве сагиттальных направлений роста.
 Эффективность управления проявляется в том, что непосредственно до, и преимущественно после гомеостазического перехода, целевая функция управления содержит компоненту форсирования процесса. Такая компонента, обычно, определяет эффективность управления на параметрах его качества, не допуская значительного перерегулирования. Этим определяется необходимая устойчивость процесса.
Эффективность управления проявляется в том, что непосредственно до, и преимущественно после гомеостазического перехода, целевая функция управления содержит компоненту форсирования процесса. Такая компонента, обычно, определяет эффективность управления на параметрах его качества, не допуская значительного перерегулирования. Этим определяется необходимая устойчивость процесса.
В то же самое время, для процессов, обусловленных парой сагиттальных представлений, отражающих непосредственные изменения NCRS ядерно-цитоплазматического индекса и прозрачности LS ядер, на интервале s [0-4], свойственно слабо форсируемое вхождение в процесс роста на уровнях состояний, соответствующих гомеостазическому переходу. Аналогичное явление имеет место и для пары инициального направления inS и прозрачности LS ядер. После гомеостазического перехода изменения на этих парах до канонизации процесса стабилизируются. Для двух других пар, нарастание форсировок после гомеостазического перехода сокращается и на конечных этапах озлокачествления, форсировки переходят в "отставания".
Аналогично, по достижению уровня роста, соответствующего 7-8 состояниям, начинается резкое снижение форсировок парой глобального и канонического сагиттальных представлений. Такой характер форсировок, по-видимому, связан с исчерпанием мощности множества клеток, трансформирующихся по этим направлениям, выполнением ими своей функциональной роли при достижении субстратом состояний уровня выше s=8 и сокращением энергоснабжения.
Таким образом, реализация форсировки имеет место по всем сагиттальным направлениям роста, что в результате их суммарного воздействия, приводит к запасам устойчивости и повышению качества управления процессом трансформации, достаточным для надежного достижения канонического состояния. Заметим, что форсирующие свойства присущи подавляющему числу известных систем управления и регуляции самого различного назначения и происхождения, в том числе – биологических.
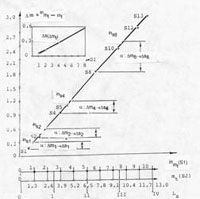 Можно говорить, что отражение форсировок равносильно отражению качества управления целевой функцией системы стабилизации неравновесного роста. Явные зависимости, отображающие форсирующие, во времени и в пространстве состояний, свойства эффекта "затягивания" в процесс роста, проявляющиеся на множествах активирующих клеток состояния PROMOTION, отражают линейность системы стабилизации неравновесного роста клеточных множеств в системе локального гомеостазиса. Это показывает рисунок, иллюстрирующий отображение форсирующих свойств множества клеток этого состояния.
Можно говорить, что отражение форсировок равносильно отражению качества управления целевой функцией системы стабилизации неравновесного роста. Явные зависимости, отображающие форсирующие, во времени и в пространстве состояний, свойства эффекта "затягивания" в процесс роста, проявляющиеся на множествах активирующих клеток состояния PROMOTION, отражают линейность системы стабилизации неравновесного роста клеточных множеств в системе локального гомеостазиса. Это показывает рисунок, иллюстрирующий отображение форсирующих свойств множества клеток этого состояния.
Линейная зависимость отображает во времени положение одноименных фаз состояния PROMOTION по отношению к положению на кривой суммарного роста множества состояний s. Так, отображенный интервал для состояния инициации указывает, что это состояние опережает усредненное по множеству сагиттальных представлений состояния s, на величину приращения s4, а разность таких интервалов на множестве {s} текущих состояний, оказывается линейной функцией. Такая функция представляется в отношении порядка фаз состояний и фазовых переходов – клеточного транспорта и характеризует функциональные свойства подмножеств клеток состояния PROMOTION как форсирующую компоненту системы локального гомеостазиса.
Так как показатели системы координат нормированы, но шкала может содержать различное число интервалов отсчета, в зависимости от типа сагиттального представления, на нижней части рисунка отражено соответствие интервалов для встречающихся, при принятой степени конкретизации, их числа равного 10 и 13, а также для сагиттального представления направления роста, отражающего парциальное влияние прозрачности ядер, для которого число интервалов отсчета, равно числу их градаций.
Линейность этой функции служит не только иллюстрацией, но и подтверждением упорядоченности роста и достоверно подтверждает сам факт управления процессом трансформации с форсировкой. Объектом этого управления является естественный технологический процесс, реализующий рост по каждому стволу и ветви древесной структуры в отдельности и по дереву в целом. Таким процессом естественной технологии является процесс организации и стабилизации групповых отношений порождения, как ответственный за агрессивное поведение клеток состояния PROMOTION и стабилизацию неравновесного состояния.
С помощью других вторичных отображений могут быть получены зависимости, отображающие более "тонкие" взаимодействия на уровне феноменологии, такие, как различные временные отображения, а также соответствующие им линейные временные диаграммы. Отображения аналогичного характера, отражают и энергетические расходы.
Древесный порядок представляет возможность отражения результативности процессов, происходящих в клетке в системе локального гомеостазиса, и отображенных в изменения обобщенного параметра озлокачествления. Такие процессы ведут к утрате клеткой функции нормальной жизнедеятельности, изменению структурной и функциональной организации клеточного множества, деградации клеток и, в результате, к их вырождению. Отражения таких изменений качества, с помощью отображений второго порядка, могут играть роль не только индикатора озлокачествления, но и численного показателя для оценки, на древесных порядках, фактора деградации биологического субстрата.
Отмечалось, что ветви детализированного дерева, с целью отображения роста между уровнями, взвешивались. В качестве веса были приняты значениях ядерно-цитоплазматического индекса таким образом, что каждому участку ветви между смежными уровнями, был присвоен вес, численно равный значению ЯЦИ, соответствующему переходу на более высокую фазу озлокачествления. Такой вес указан на ветвях дерева, и он “взвешивает” нарастающее изменение состояния множества клеток при переходе на следующий уровень. Счетно-аддитивная функция, отображающая процесс накопления весов, есть отображение деградации множества N клеток, образующих древесный порядок. Она обладает всеми свойствами, рассмотренными ранее для отображения сложности дерева в пространство состояний.
Это означает, что формально, под деградацией множества клеток, отображенной на древесный порядок, понимается накопление в пространстве состояний и во времени, значений ядерно-цитоплазматического индекса, отраженного в рост мощности множества клеток обобщенного параметра озлокачествления, образующего по уровням состояний древесного роста, совершенный строгий порядок. Полагается, как и ранее, что время выступает в качестве параметра отображения.
Деградация отображается в виде нарастающей вдоль ствола дерева, по мере увеличения степени поражения субстрата, последовательной суммы деградации на его уровнях. Деградация на уровне определяется суммой весов межуровневых участков ветвей, образующих уровень, деградация на страте состояния определяется суммированием веса ветвей на межстратовых уровнях.
Таким образом, отображение деградации на древесном росте, отражая дегенерацию субстрата, формируется с учетом разрастания дерева вдоль уровней в функции значения (имени) уровня озлокачествления, идентифицируемого именем текущего состояния. Это означает, что отображение деградации образуется с учетом разрастания дерева вдоль уровней в функции значения (имени) уровня озлокачествления, идентифицируемого именем текущего состояния.
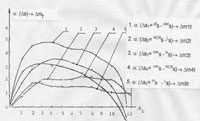
На титульном рисунке приведены зависимости, иллюстрирующие отображение фактора деградации субстрата, принадлежащего состояниям PROGRESSION и PROMOTION. На нем показаны отображения дерева деградации клеток биологического субстрата и его парциальных составляющих – нормированный и суммарный показатели дегенерации; деградация множества клеток состояния PROMOTION с учетом агрессивности типа “возбуждения”; собственная деградация множества клеток состояния PROMOTION (без учета агрессивности) и суммарная деградация состояния PROGRESSION. Буквами латинского алфавита A, B, C,…F обозначены идентификаторы ветвей кроны поддерева PROGRESSION.
Сигнал ошибки управления неравновесным состоянием, как входной сигнал системы с обратной связью, отражен разностью деградации, заданной целевой функцией управления системы локального гомеостазиса и деградации, осуществимой в текущем состоянии. Явно отображен резкий рост деградации, порожденный множеством клеток баланса эквивалентности на этапе гомеостазического перехода из инициального состояния s=P4, PG0. Это четко отражает ролевую функцию этого множества. Резкое увеличение деградации имеет характер скачка, и существенно опережает деградацию клеток состояния PROMOTION, порожденного их активационной агрессивностью. Можно говорить, что это есть отражение функции форсировки, как «штурмовой» функции активации.
Видно, что агрессивность типа “возбуждения” на 3-5 уровнях форсирует дегенеративные изменения. Достижение суммарной деградации голыми ядрами BN реализуется полной отработкой управляющего сигнала ошибки системы стабилизации роста.
Поэтому можно считать, что целевая функция локального гомеостазиса осуществляет задачу достижения деградационными процессами дегенеративного состояния субстрата с параметрами, отраженными на его вторичных отображениях.
Аналогичные построения могут быть произведены и в пространство времени, а также и для каждой отдельной ветви структуры. В частности, будучи построены в естественных единицах для пространства PROGRЕSSION, они отобразили отношение порядка деградации множеств клеток по уровням процесса озлокачествления в виде "колосящегося" дерева.
Отношение порядка деградации клеточного множества на множестве уровней состояния древесной структуры, отражается отношением предпочтения, определяемого путями роста множеств озлокачествленных клеток, доставляющими оптимальный, в смысле максимальной деградации, древесный рост.
Такое отношение предпочтения можно отразить отношением упорядочения на ветвях древесного порядка, выраженным в виде порядка множеств обобщенного параметра озлокачествления, идентифицированных номерами его ответвлений. При этом, на некотором множестве обобщенных параметров состояния PROMOTION, между отношениями предпочтения устанавливается отношение эквивалентности. Пространство, образующее множество парциальных отображений деградации образует подпространство деградации фазового пространства состояний.
Яков Гельфандбейн
© 1011494 CIPO



Популярное
Mazda и Toyota переиграли рынок: один завод в США стал щитом от новых пошлин. Рассказываем, как им удалось просчитать ситуацию на годы вперёд.



Последние материалы

Эл № ФС77-72263 выдано РКН 01.02.2018. Учредитель: В.В. Горшенин. Главный редактор: И.С. Новикова.
Контакты: +7 (499) 641-41-69, 105066, а/я №26 (для официальных обращений), home@pravda.ru Москва, ул. Старая Басманная, д.16/1Б, «Правда.Ру»
Copyright © 1999-2025, технология и дизайн принадлежат ООО «Техномедиа».
Материалы сайта предназначены для лиц старше 18 лет (18+).
*Meta Platforms признана экстремистской организацией и запрещена, как и принадлежащие ей Facebook и Instagram.